40. petrology and carbon and oxygen stable isotopic
TRANSCRIPT

40. PETROLOGY AND CARBON AND OXYGEN STABLE ISOTOPIC COMPOSITION OFMACROFOSSILS AND SEDIMENTS FROM THE BLAKE-BAHAMA FORMATION,
DEEP SEA DRILLING PROJECT SITE 603, WESTERN NORTH ATLANTICLOWER CONTINENTAL RISE1
Janet A. Haggerty, Department of Geosciences, University of Tulsa2
ABSTRACT
Strata that record the evolutionary history of the North American continental margin in a region that serves as thebasin margin interface between allochthonous sedimentation from the continent and pelagic sedimentation from theoceanic realm were recovered at Deep Sea Drilling Project Site 603, on the lower continental rise. The lowermost unitrecovered at this site is composed of upper Berriasian-Aptian interbedded laminated limestone and bioturbated lime-stone with sandstone to claystone turbidites. This unit can be correlated with the Blake-Bahama Formation in the west-ern North Atlantic. Studies of the laminated and bioturbated limestones were used to determine the depositional envi-ronment. Geochemical and petrographic studies suggest that the laminated limestones were deposited from the suspend-ed particulate loads of the nepheloid layer associated with weak bottom-current activity as well as moderate to poorlyoxygenated bottom-water conditions.
Fragments of macrofossils are also found in the Blake-Bahama Formation drilled at Site 603. Twelve specimens andtheir host sediment were analyzed for their carbon and oxygen isotopic composition. The macrofossil samples chosenfor analysis consist of nine samples of Inoceramus, two ammonite aptychi, and one belemnite sample. Depletion in 18Ois observed in recrystallized specimens. The ammonite aptychi have been diagenetically altered and/or exhibit evidenceof isotopic fractionation by the organism. Oxygen isotope paleotemperatures obtained from five well-preserved speci-mens—four of Inoceramus and one of a belemnite—suggest that bottom-water temperatures in the North Atlantic Ba-sin during the Early Cretaceous were very warm, at least 11°C.
INTRODUCTION
Hole 603B, drilled on DSDP Leg 93, penetrated 1585 mof section. This is the first deeply buried sediment re-covered from beneath the lower continental rise of thewestern North Atlantic Basin. The site is located in 4633 mwater depth, approximately 435 km from Cape Hatteras,North Carolina (Fig. 1). Site 603 is located adjacent tothe lower continental rise terrace in the first valley of thelower continental rise hills. The position of the site, andimprovements in drilling technology since the early legsof the Deep Sea Drilling Project, make drilling results atthis site uniquely different from the drilling results atprevious DSDP sites in the western North Atlantic. Stratarecovered at Site 603 record the evolutionary history ofthe continental margin in a region functioning as the ba-sin margin interface between allochthonous sedimenta-tion from the continent and pelagic sedimentation fromthe oceanic realm.
The sedimentary section recovered at Site 603 is di-vided into five lithologic units (I through V; Lithologysection in Site 603 chapter, this volume). These five unitscan be correlated with the oceanic formations proposedfor the western North American Basin by Jansa et al.(1979). The deepest unit (Unit V) is composed of 261 mof upper Berriasian-Aptian interbedded laminated lime-stone and bioturbated limestone with sandstone to clay-
1 van Hinte, J. E., Wise, S. W., Jr., et al., Init. RepCs. DSDP, 93: Washington (US.Govt. Printing Office).
2 Address: Dept. of Geosciences, University of Tulsa, 600 South College Avenue, Tulsa,OK 74104.
stone turbidites. This unit is the local equivalent of theupper portion of the Blake-Bahama Formation as de-fined by Jansa et al. (1979) from the type locality atDSDP Site 391 (Benson, Sheridan, et al., 1978).
The author uses the term "laminated limestone" rath-er than "laminated marl" as listed in the Site 603 chap-ter (this volume), because the carbonate content in theserocks is usually greater than 60%. The term "marl" isused for rocks that contain 30 to 60% carbonate, withthe remaining components being mostly clays (see Ex-planatory Notes, this volume).
The sedimentologists on board Leg 93 divided Unit Vinto two subunits on the basis of the presence or ab-sence of turbidites. Subunit VB, upper Valanginian toupper Berriasian, lacks turbidites. Subunit VA, Aptianto upper Valanginian, contains numerous turbidites. Thepresence of terrigenous clastic turbidites in Subunit VAmakes the Blake-Bahama Formation at Site 603 differ-ent from other DSDP sites. The Blake-Bahama Forma-tion is characterized by cycles of laminated marlstone orlimestone and bioturbated limestone (Fig. 2). Anotherinteresting feature in the Blake-Bahama limestones ofUnit V at Site 603, is the inclusion of fragments of mac-rofossils. Macrofossils are not abundant in these lime-stones, but their occurrence is usually associated withthe laminated limestone; they are rarely found in the bio-turbated limestone.
The purpose of this chapter is to investigate the depo-sitional environment of the limestones of the Blake-Ba-hama Formation, and the carbon and oxygen stable iso-topic composition of the limestones and their macrofos-sils, with the goal of elucidating the early history of this
1003

J. A. HAGGERTY
45°N
4 0 ;
"I 1 1 1 1 1 1 f
\S hVTTTTTT>^^<rT-rTTTTTTT=^—-
7 f Sea A
ε^M K J? I I I 13
85°W
Figure 1. Location map with physiographic features of the North American Basin and location of DSDP sites(from Legs 2, 11, 43, 44, 76, and 93). Site 603 is located in 4633 m water depth on the lower continental rise.Bathymetry after Uchupi (1971).
restricted, narrow, and youthful North Atlantic Oceanbasin. Determination of oxygen isotope paleotempera-tures can provide valuable data for understanding thepaleoenvironment of the Early Cretaceous North Atlan-tic.
A vast number of paleotemperature estimates havebeen made on outcrops of Cretaceous strata that were
deposited in the Cretaceous epicontinental seas. Severalstudies have analyzed the isotopic composition of Ino-cerαmus and belemnites from these shallow sea depos-its (Lowenstam and Epstein, 1954; Stevens and Clayton,1971). Skeletons from these organisms were chosen foranalysis because they are not as susceptible to diageneticalteration as many other skeletal remains. However, their
1004

PETROLOGY AND CARBON AND OXYGEN STABLE ISOTOPIC COMPOSITION
0 r79-1 79-2 79-3 79-4 79-5 79-6 79-7
10
20
30
40
50
60
70
80
90
100
110
120 -
130 -
140,-
150*-
Figure 2. Typical core from the Blake-Bahama Formation. Note thecharacteristic alternations of laminated limestone with bioturbatedlimestone. The core is from Subunit VB, therefore clastic terrige-nous turbidites are not present. This core (603B-79) contains frag-ments of macrofossils in the laminated limestone.
coarse crystalline nature and calcite mineralogic compo-sition is not always sufficiently resistant to diagenetic al-teration from subaerial exposure and meteoric diagene-sis. This makes data derived from outcrop samples verydifficult to interpret.
Macrofossils deposited in ancient deep-sea sedimentsthat have not been uplifted on land usually have been
subjected to only burial diagenesis, Extension of conti-nentally derived fresh-water aquifers into marine sedi-ments has only been recorded in near-continent hemipe-lagic sediments that have layers of permeable sandstone(Gieskes, 1981). From the interstitial water analyses per-formed on board ship, there is no evidence of fresh-wa-ter aquifer penetration into the pelagic sediments of UnitV at Site 603 (Site 603 chapter, this volume).
Very few studies have been conducted on macrofos-sils from deep sea cores. Only Barron et al. (1984) andSaltzman and Barron (1982) have reported oxygen iso-tope paleotemperatures from Inoceramus extracted fromDSDP cores. After a rigorous selection procedure, Saltz-man and Barron (1982) used only 49 specimens for de-termination of oxygen isotope paleotemperatures froman initial group of 500 Inoceramus specimens from DSDPcores. These 49 specimens—less than 10% of the totalnumber of specimens—were deemed well preserved. Ex-cellent preservation of Inoceramus specimens for paleo-temperature determinations is not common in deep-seacore material.
In this study, Inoceramus, ammonite aptychi, a bel-emnite, and the host sediment have been analyzed forcarbon and oxygen isotopes. Several types of macrofos-sils were chosen for isotopic analysis to see if the resultsfrom different taxa would yield distinctly different iso-topic data.
DISTRIBUTION OF MACROFOSSILS IN CORESFROM HOLE 603B
A few fossil ammonites (Fig. 3A), as well as ammo-nite aptychi (Fig. 3B), Inoceramus, and shallow-watercarbonate debris are incorporated in the sediment of UnitV. Table 1 lists the sample and the host lithology foreach occurrence; the data are from the shipboard visualcore descriptions and additional observations made bythe author. Visual examination was limited to the twofaces of the split core, therefore some macro fossil mate-rial may not have been detected.
The shallow-water carbonate debris is a minor to tracecomponent contained within the terrigenous sandstoneturbidites in Subunit VA. Some fragments of inverte-brate remains that can be identified to the general taxalevel are gastropods, pelecypods, and bryozoans (Bio-stratigraphy section in the Site 603 chapter, this volume).The other macroinvertebrate debris is usually shell frag-ments that are small and abraded, lacking characteristicfeatures for additional identification. They do not ap-pear to be fragments from delicate skeletal remains ofaragonitic ammonites or calcareous Inoceramus shells.The carbonate debris is interpreted as being derived fromshallow water because of the association with rare andpoorly preserved shallow-water, midshelf benthic fora-minifers (Biostratigraphy section in Site 603 chapter, thisvolume). The shallow-water skeletal debris and the abun-dant terrigenous elastics were transported from the shelfand redeposited in these turbidites associated with thesubmarine fan facies (see Sarti and von Rad, this vol-ume, for a detailed discussion of the submarine fan fa-cies).
1005

1 cm
1 cm
Figure 3. A. A Lower Cretaceous (Barremian-Aptian) ammonite on the bedding plane surface of the full-round of the core (Sample603B-44-3, 140 cm). See Frontispiece Figure D (this volume) for a color photograph of this ammonite, and Hoedemaker (this volume) forpaleontologic details of the Lower Cretaceous ammonites. B. An aptychus found on the bedding plane of a quarter-round of the core.Upper and lower bedding plane surface shown. Note the dark chitinous layer on the surface of the aptychus. This sample (603B-79-2,94.5 cm) was found in the core shown in Figure 2.
The majority of the remaining macro fossils are im-pressions of flattened ammonites, calcite ammonite ap-tychi, or Inoceramus. These are predominantly found inthe laminated limestone associated with Subunit VB(Fig. 2). A rare fragment of a belemnite (Fig. 4A) is alsonoted in the bioturbated limestone of Subunit VB. SeeHoedemaker (this volume) for details on the taxonomyand biostratigraphic significance of the Early Cretaceousammonites.
METHODS
After surveying the samples listed in Table 1, the author made a se-lection of samples that included Inoceramus, ammonite aptychi, anda portion of the belemnite rostrum fragment, as well as the host rockfor isotopic analysis. No shallow-water carbonate debris were selectedfor analysis. DSDP policy does not permit researchers to decimate thecore by splitting it on the bedding plane to find macrofossils or to de-termine the size or degree of preservation of the macrofossil fragmentsduring investigations on board ship or at the core repository. There-fore, samples were chosen based upon examination of the cut face of
1006

PETROLOGY AND CARBON AND OXYGEN STABLE ISOTOPIC COMPOSITION
Table 1. Distribution of macrofossils in cores from Hole603B.
Core-Section,interval orlevel in cm Lithology Macrofossil
44-3, 15050-1, 124-12650-1, 0-8052-5, 12-12253-3, 52-8056-1, 120-12557-2, 59-6358-161-463-2, 61-6563-3, 10-2069-5, 15-2075-3, 113-11775-3, 131
Laminated limestoneLaminated limestoneSandstoneSandstoneSandstoneSandstoneSandstoneLaminated limestoneLaminated limestoneBioturbated limestoneLaminated limestoneLaminated limestoneLaminated limestoneLaminated limestone
AmmoniteInoceramus, aptychusShell debrisShell debrisShell debrisShell debrisShell debrisAptychusAptychiInoceramusAptychusAmmoniteAptychiAptychus
76-1, 120 Base of Subunit VA
76-3, 7177-1, 48-5277-3, 48-5278-1, 51-5278-2, 9478-2, 11078-3, 65-6679-1, 2179-1, 3679-1, 10979-2, 58-5979-2, 9479-3, 70-12079-579-5, 16-2079-6, 56-5779-6, 84-9680-1, 6-780-2, 125-13080-2, 145-15080-3, 20-2180-3, 34-3680-4, 60-6581-5, 15-17
Laminated limestoneBioturbated limestoneLaminated limestoneLaminated limestoneLaminated limestoneLaminated limestoneLaminated limestoneLaminated limestoneLaminated limestoneLaminated limestoneLaminated limestoneLaminated limestoneLaminated limestoneLaminated limestoneLaminated limestoneLaminated limestoneLaminated limestoneLaminated limestoneLaminated limestoneLaminated limestoneLaminated limestoneLaminated limestoneLaminated limestoneLaminated limestone
AptychusBelemniteInoceramusInoceramus, aptychiAmmoniteAmmoniteInoceramusAptychusAptychusAptychiAptychiAptychusAptychiAptychiAptychiInoceramus, aptychiAptychi, InoceramusInoceramus, aptychiInoceramusInoceramusInoceramusInoceramusInoceramusInoceramus
the working half of the cores. (Sampling was not permitted of the ar-chived half of the cores.) During shore-based studies, the samples weresplit along bedding planes to determine the size of the macrofossilfragments and to study the degree of alteration of the shell. Many ofthe fragments were too small for thin-section preparation with the hostsediment. Thin sections containing the macrofossils and their immedi-ate host rock were unsuitable for additional geochemical analysis byelectron microprobe. The abundance of clay in the host rock made thethin-section preparation very difficult, and the sample could not bepolished for quantitative microprobe analysis.
All hand specimens were examined with the use of a stereomicro-scope for evidence of diagenesis. Subsamples of the carbonate shellwere taken from the belemnite fragment, and aptychi fragments bybreaking off pieces and carefully cleaning the exterior of any host sed-iment or organic chitinous coating (see Fig. 4B). Samples were takenfrom the calcite prismatic layer of Inoceramus fragments by breakingoff pieces or by extracting individual crystals with a picking brush.Bulk rock samples of the host sediment were subsampled at the sametime for isotopic analysis.
Samples were prepared for thin sectioning after they were subsam-pled for isotopic analysis. The rocks were pressure impregnated withepoxy resin and then cut and ground for thin section.
The macrofossil samples and the host rock samples for isotopicanalysis were vacuum roasted at 35O°C for an hour to remove volatilecomponents. Carbon and oxygen stable isotopic compositions of thecarbonate components of the bulk rock and of the macrofossils weremeasured by analyzing the carbon dioxide generated from the reactionof the samples with 100% phosphoric acid—a standard technique es-tablished by McCrea (1950). The carbon dioxide gas sample was ana-
Figure 4. A. Photomicrograph (under crossed nicols; bar scale = 1 mm)of a cross section of a belemnite in bioturbated limestone (Sample603B-77-1, 48-52 cm). The belemnite has been infilled with sparrycalcite but preservation of the shell is excellent; note the presenceof pseudoisogyres in the shell. B. Chitinous organic coating on ap-tychus (Sample 603B-79-2, 58-59 cm; reflected light; bar scale =1 mm).
lyzed on a Finnigan MAT 251, an isotope ratio mass spectrometer.Analyses are reported relative to PDB (Pee Dee Belemnite) standard.Analytical error was no greater than ±O.2%o.
The aptychi of the ammonites, the rostrum of the belemnite, andthe prismatic layer of Inoceramus shells were originally composed ofcalcite (Majewske, 1974). Therefore, the remains of the pelagic macro-fossils, and epibenthic Inoceramus were chosen for isotopic analysisbecause they are more likely to have survived diagenesis without re-crystallization and isotopic re-equilibration. The purpose of analyzingthe isotopic composition of these skeletal remains was to obtain theoxygen isotope paleotemperatures for the Early Cretaceous in the west-ern North Atlantic. This approach to acquiring paleotemperatures isnot new and has previously provided valuable information about deep-sea circulation in the Late Cretaceous (Saltzman and Barron, 1982).
The estimation of paleotemperatures from the oxygen isotopic com-position of the shell assumes that the shell was biogenically precipi-tated in isotopic equilibrium with the surrounding seawater when theorganism was alive. Changes in the oxygen isotopic composition ofthe shell should therefore reflect changes in the temperature of the sea-water, assuming no complications have arisen with glaciation or localdifferences in salinity. The original isotopic composition of the shellmust be retained unaltered after the death of the organism for properacquisition of an oxygen isotope paleotemperature.
1007

J. A. HAGGERTY
Many organisms do not necessarily precipitate their shells in iso-topic equilibrium with the surrounding seawater, but impart a "vitaleffect" or shift in the oxygen isotopic signature of the shell (e.g., We-ber and Woodhead, 1972). Yet several studies have analyzed the iso-topic composition of Inoceramus and belemnites from Cretaceous epi-continental seas (Lowenstam and Epstein, 1954; Stevens and Clayton,1971) and Inoceramus from deep-sea sediments (Barron et al., 1984,Saltzman and Barron, 1982) and assumed that the shells were secretedin isotopic equilibrium with the surrounding seawater. The oxygen iso-topic composition of ammonite aptychi have not been previously re-ported in the literature for estimating paleotemperatures, and carefulevaluation is required before assuming skeletal precipitation in isotop-ic equilibrium with seawater.
The method of calculating paleotemperatures of the Cretaceousseawater from the oxygen isotopic composition of Inoceramus thatwas employed by Saltzman and Barron (1982) is used in this study.This method makes use of an Epstein paleotemperature equation pub-lished by Yapp (1979):
7X°C) = 16.4 - 4.2(δc - δw) + 0.13(δc - δ w ) 2
where δc = δ 1 8 θ (PDB) of the calcium carbonate, and δ w = δ 1 8 θ(SMOW) of the water.
The calculations performed in this study, and in Saltzman and Bar-ron (1982), assume that the oxygen isotopic composition of mean oceanwater during the Cretaceous was depleted by one per mil relative to theSMOW (Standard Mean Ocean Water) standard. This depletion of oneper mil is used for the oxygen isotopic composition of seawater duringthe Cretaceous, because it is assumed that the polar ice volume wasnegligible during the Cretaceous (Shackleton and Kennett, 1975). Thechoice of this value for the water correction factor and this paleotem-perature equation may be debated, but for the purposes of comparingdata from this study with that of the Saltzman and Barron (1982)study, this value and paleotemperature equation are appropriate.
RESULTS
Petrology
The Blake-Bahama Formation correlates with Unit Vat Site 603. This upper Berriasian to Aptian unit is com-posed of interbedded laminated limestone and bioturbat-ed limestone with sandstone and clay stone turbidites, andis divided into two subunits on the basis of the presenceor absence of sandstone or claystone turbidites. The baseof Subunit VA is placed at the base of the lowest clay-stone turbidite, Sample 603B-76-1, 120 cm. The lami-nated limestone and the bioturbated limestone are verysimilar throughout the two subunits, and therefore thedescriptions of the two rock types are representative ofthe whole unit. In the Site 603 chapter (this volume), thelaminated limestone and the bioturbated limestone arefrequently referred to as in situ pelagic sediments. Thisinterpretation should be re-evaluated in light of the re-sults of this study, which describe the variations and dif-ferences of the limestones at DSDP Site 603.
The bioturbated and laminated limestones can be dis-tinguished on the basis of their biologic sedimentary anddepositional sedimentary structures. The two basic endmembers are distinctly laminated or bioturbated, but inseveral locations a transitional form is observed in whichbioturbation has not completely destroyed the thin lami-nations (Fig. 5A). Any of these types of limestones mayabut against an overlying turbidite (Fig. 5B). The per-cent of the core containing laminated limestones or marlsincreases from the bottom of Unit V to the base of Sub-unit VA where the claystone turbidites appear (see thecore photos at the end of the Site 603 chapter, this vol-
105 i—
110
115
120
125
130
1 3 5 1 -
6 0 1 -
65
70
90
Figure 5. A. Bioturbated limestone interbedded with laminated lime-stone (Subunit VB, Sample 603B-74-4, 105-135 cm). Note the darkcoloration of the clay-rich burrow fills. B. Typical laminated lime-stone and bioturbated limestone of the Blake-Bahama Formationfound in Unit V at Site 603 (Sample 603B-70-3, 60-90 cm). Noteareas in the cores that are an intermediate form, that is, indicatingcharacteristics of bioturbation and faint lamination. This core isfrom Subunit VA and therefore contains deposits of turbidites, vis-ible at the very upper and lower edges of the photographed interval.
ume). An inverse trend in the percent of core displayingevidence of bioturbation is also apparent in Subunit VB.
The variable amounts of clay, organic matter, and cal-cium carbonate contained within the limestones is di-rectly related to their numerically assigned color usingthe Soil Color Charts of the Munsell Color Company.The darker colors (lower numbers) have a higher per-centage of clay and/or organic matter, or a lower per-centage of calcium carbonate. In some portions of thelaminated lithology, the sediment is sufficiently clay richto be termed a marl or marlstone (depending on degreeof induration).
A comparison of the shipboard carbonate bomb dataand organic carbon data of these three types of lime-
1008

PETROLOGY AND CARBON AND OXYGEN STABLE ISOTOPIC COMPOSITION
stone—bioturbated, laminated, and transitional (lami-nated and bioturbated)—displays some interesting dif-ferences (see Table 2). The laminated limestone ranges incolor from dark gray (N2, 5Y 2/1) to medium gray (N6,5Y 6/1) with a brownish tint, and has an average car-bonate content of 68% and an organic carbon contentof 1.41%. The bioturbated limestone ranges in colorfrom medium gray (N5) to nearly white (N8), and has anaverage carbonate content of 78% and an average or-ganic carbon content of 0.19%. The transitional lime-stone, which shows characteristics of being laminatedand bioturbated, ranges in color from medium gray (N4)to light gray (N7), and has intermediate values of 69%for the average carbonate content and 0.33% for the av-erage organic carbon content. The direction of the vari-ations in the average carbonate and organic carbon con-tent percentages agrees with the prediction based uponcolor of the sediment.
A significant trend that is suggested from the valueslisted in Table 2 is an increase in both the carbonate con-tent and the organic carbon content from the lower Sub-unit VB to the upper Subunit VA of Hole 603B. There isa wide range and overlap in the values as shown by thestandard deviation, but this trend is distinctly representedin the mean value for each of the three types of lime-stone. This trend is not a function of the color of thelimestone because a detailed dissection of the data bycolor and type of limestone shows the same trend be-tween Subunit VB and Subunit VA.
The bioturbated clay-rich nannofossil limestone con-tains abundant calcified or pyritized radiolarians. NoInoceramus fragments or ammonite aptychi have beenfound in this type of sediment. One belemnite fragmentis found in Sample 603B-77-1, 48-52 cm (Fig. 4A). Thebioturbation is detected in thin section by the patternsgenerated from either the increased concentrations of ra-diolarians within burrows and the patchiness of the mi-critic matrix (Fig. 6A) or by the color and textural dif-ference of the increased clay content in a burrow (Fig.5). Radiolarians, as well as burrows, are sometimes flat-tened and oriented parallel to bedding, therefore indi-cating compaction. Thin sections of some samples, de-noted as bioturbated limestone in the Site 603 chapter(this volume), show layers that are enriched in radiolari-ans (Fig. 6B and C), but the hand samples display nocolor change indicative of a burrow. The enrichment of
Table 2. Carbonate and organic carbon content of "pelagic lime-stones" from Unit V of Hole 603B.
CarbonateTexture Subunit
Organic carbon(ft)
No. ofanalyses
Laminated limestones
Bioturbated limestones
Bioturbated andlaminated lime-stones
VAVBV
VAVBV
VAVBV
696368
797278
706569
±±±
±±±
±±±
18517
5158
8109
1.441.271.41
0.210.070.19
0.390.110.33
±±±
±±±
±±±
0.60.50.6
0.10.070.14
0.280.030.27
286
34
204
24
133
16
radiolarians in these layers may be interpreted as the re-sult of bioturbation concentrating the radiolarians with-out an increase in the clay and/or organic content of theburrow or as evidence of deposition associated with anincrease in the supply of radiolarians or very weak cur-rent activity.
The ichnofabric in the bioturbated limestone displaysa distinct decrease in burrow size from the bottom ofthe white limestone to the transitional bioturbated andlaminated limestone (Figs. 5 and 7). This decrease inburrow size is easily observed because of the darker col-or of the burrow fill. Tiering relationships of the infaunaare observed whereby Planolites is crosscut by Chon-drites. Dark transitional limestone, with a laminated sed-imentary fabric that contains evidence of bioturbation,is usually associated with very small-diameter burrows(Fig. 5); larger-diameter burrows are rarely observed inthe dark transitional limestones unless they are withinapproximately 2 cm of an overlying light gray limestone.
The transitional form of a bioturbated, faintly lami-nated, clay-rich nannofossil limestone (Fig. 7A) may oc-cur between two end members of bioturbated or lami-nated limestone, or one end member may not be present(Fig. 7B). Only one occurrence of a macrofossil in thistype of transitional sediment is noted from Hole 603B(Sample 603B-81-5, 15-17 cm; Fig. 7A). Figures 6C, 8A,and 8B show the sequence of change over an 8-cm in-terval in Core 603B-75 from Subunit VB. Figure 6C isfrom bioturbated limestone in which radiolarians appearto be flattened from compaction. In Figure 8A the radi-olarians are slightly concentrated in layers from currentaction. Figure 8B, which completes the sequence, showsthat there has been a major decrease in the concentra-tion of radiolarians and an increase in the clay content.
A microlenticular structure appears in the laminatedlimestone. The source of these lenticular structures maybe diagenetic concentrations of silica and/or slight butsignificant changes in the constituent grains during de-position of the sediment. In thin section, these struc-tures are resolved to be microlenticularlike bedding withsingle (isolated) lenses in what appears in hand speci-men to be the distinctly laminated limestone (Fig. 9).The majority of these lenses are discontinuous and donot contain as much clay and organic debris as does thesurrounding sediment. The lenses are exceptionally thin,giving a laminated appearance to hand samples of therock. The outline of some lenses is enhanced by com-paction of the clay and organic debris. The laminatedlimestone contains a terrigenous silt component of fine-grained mica, and also land plant debris as exhibited bywood fragments. These lenses or laminations (< 1 mmin thickness) indicate that some bottom currents wereactive during deposition of the sediment. The appear-ance of the specimen is reminiscent of "Lithofacies G"of Mutti and Ricci Lucchi (1972).
The majority of the macro fossils found at Site 603are deposited in the laminated sediment (Fig. 10). Thedegree of preservation of the macrofossils is variable.Some specimens are partially pyritized, others exhibitpartial to complete recrystallization of the calcite, andothers appear to be well preserved.
1009

J. A. HAGGERTY
A
* • • • ' • . \ . * • > • # • .
Figure 6. A. Bioturbated limestone of the Blake-Bahama Formation (Sample 603B-77-2, 42-46 cm). Concentrations of calcified radiolarians are in-corporated in burrow fills (plane polarized light; bar scale = 1 mm). B. Bioturbated limestone containing a layer with abundant calcified radio-larians (Sample 603B-82-2, 148-150 cm). These layers are probably the result of current activity winnowing away some of the fine clays and mi-crite (plane polarized light; bar scale = 0.5 mm). C. Bioturbated limestone of the Blake-Bahama Formation (Sample 603B-75-5, 1-8 cm) withpatchy distribution of calcified and flattened radiolarians; flattening is probably the result of compaction (plane polarized light; bar scale =0.5 mm).
Close inspection of the contact between the macro-fossils and the laminated sediment (Fig. 11) reveals noevidence of scouring activity around the fossil. Any dis-tortion in the laminations appears to be the result ofcompaction of the soft sediment against the morpholog-ic relief of the shell. On the basis of these observations,the currents that existed to develop the microlenticularbedding or laminations are interpreted to have been in-capable of overturning or scouring around the macro-fossils.
Inspection of one of the bedding planes in the dis-tinctly laminated limestone-containing macrofossils (603B-
78-3, 65-66 cm) reveals disarticulated skeletal calciteprisms (Fig. 12A) associated with bioturbation (Fig. 12B)and boring of the shell (Fig. 12C). Mirror images of thetop (Fig. 12D) and bottom (Fig. 12E) bedding planesshow that boring of the shell by infaunal organisms scat-tered the prisms in the immediate overlying sediment.The existence of infaunal organisms are evidence of atleast a moderately oxygenated environment.
Radiolarians are not abundant in the laminated sedi-ment. Foraminifers are very rare and have a sporadic oc-currence in these laminated sediments. The low diversityof foraminiferal assemblages and the small size of the
1010

PETROLOGY AND CARBON AND OXYGEN STABLE ISOTOPIC COMPOSITION
90
95
100
105
110
115
120 -
Figure 7. A. Transitional limestone (Sample 603B-81-5, 15-17 cm); bioturbation has not destroyed faintlaminations. Note the rare occurrence of a shell from a macro fossil incorporated in the bioturbatedsediment; the burrow is slightly flattened from compaction. Also note the dark clay-rich infilling ofthe burrow (reflected light; bar scale = 1 mm). B. Transitional limestone (with characteristics of bio-turbation and faint lamination), associated with two cycles of laminated limestone (Sample 603B-81-2,90-120 cm). Note that flattening of the dark burrows enhances the appearance of the laminations.
benthic foraminifers is suggested to be indicative of adeep-water calcareous and agglutinated assemblage thatmay have been transported from their original biotope(Biostratigraphy section in the Site 603 chapter, this vol-ume). The calcium carbonate content (see Table 2) and
the darker coloration of the sediment in comparison tothe bioturbated limestone suggest that nannofossils areprobably not as abundant as in the latter. The combina-tion of the above-described observations of (1) the exist-ence of infaunal organisms, accompanied by (2) a de-
1011

Figure 8. A. Second in a series displaying the transition from bioturbated limestone to laminated limestone in the Blake-Bahama Formation (Sample6O3B-75-5, 1-8 cm). (Fig. 6C is the first in the series.) The distribution of radiolarians is concentrated into layers, and their abundance is decreas-ing (plane polarized light; bar scale = 0.5 mm). B. The last of a series displaying the transition from bioturbated limestone to laminated lime-stone (Sample 603B-75-5, 1-8 cm). (The previous two photographs in the sequence are Figs. 6C and 8A.) The transition is marked by the changein color and appearance of microlenticularlike structures (plane polarized light; bar scale = 0.5 mm).
Figure 9. Laminated limestone from Blake-Bahama Formation at Site603, which contains microlenticularlike structures whose outline isenhanced by compaction of surrounding clays (Sample 603B-77-3,48-52 cm). The lenses may be a product of diagenesis but probablyare related to a difference in the original sediment that was depos-ited (plane polarized light; bar scale = 0.5 mm).
crease in bioturbation activity and burrow size, as wellas (3) an increase in terrigenous clastic components, and(4) a decrease in the abundance of microfossils in thelaminated sediments indicates that the rate of deposi-tion of the laminated sediments may have been greaterthan that of the bioturbated limestone. The laminatedstrata are not interpreted as an anaerobic facies becauseof the presence of epibenthic macrofossils and evidenceof infaunal activity.
Stable IsotopesTable 3 lists the type of macrofossil, the stable isotop-
ic composition of the macrofossil, and that of the hostrock. The lithology of the host rock for all of the Inoce-ramus and aptychi listed in Table 3 is laminated lime-stone; only the belemnite was contained in bioturbatedlimestone. This table also contains data for additionalsamples of limestone from Prezbindowski and Pittman(this volume); their samples of limestone were pretreat-ed by the same method as the samples in this study andwere analyzed in the same laboratory. Therefore, it isreasonable to combine the two data sets.
1012

PETROLOGY AND CARBON AND OXYGEN STABLE ISOTOPIC COMPOSITION
83
84
85
86
87
88
89
90
91
92
93
94
95
96
97 -
98 "―
Figure 10. Cut surface of core displaying fragments of aptychi andInoceramus contained in the laminated limestone (Sample 6O3B-79-6, 83-98 cm). (Close-up of core shown in Figure 2.) Note theorientation of the aptychus located at 90 cm.
Figure 13 is a cross plot of the data listed in Table 3.The plot shows a scatter of points from all lithologiesand macro fossils. There does not appear to be any trendwith increasing sub-bottom depth. Two statements can
Figure 11. Laminated limestone with macrofossil (Sample 603B-78-1,51-52 cm). There is no evidence of current scouring around thefossil; the only distortion of the laminations is from compaction ofsediment (reflected light; bar scale 1 mm).
be made from an evaluation of this plot. The aptychiare significantly depleted in both 1 3C and 1 8O by com-parison with the other in samples, and the bioturbatedlimestone also appears to be slightly depleted in 1 8O rel-ative to the laminated limestone and Inoceramus frag-ments. No definitive statements can be made as to wheth-er there is a pronounced enrichment in 1 3C in either thebioturbated or laminated limestones or even if there isan enrichment in 1 3C in the limestones from Subunit VAto those of Subunit VB.
Diagenetic Trends
In order to evaluate the effect of diagenesis on theisotopic composition of Inoceramus, two specimens (Sam-ples 603B-78-1, 51-58 cm and 603B-80-3, 20-21 cm) thatclearly exhibited the effects of recrystallization in theprismatic layer were analyzed. Sample 6O3B-8O-3, 20-21cm had more than one specimen of Inoceramus avail-able for subsampling; one of these specimens displayeda cloudy white coloration in the prisms, indicating thatrecrystallization had occurred. The two specimens thatdistinctly exhibited diagenetic alteration yielded valuesthat were depleted in 1 8O (-3.3 and -3.5%o) and 1 3C(-0.3 and O.2%o) relative to the isotopic composition ofthe well-preserved specimen from Sample 603B-80-3,20-21 cm (see Table 3). Barron et al. (1984) reported val-ues as light as - 6.75%o in δ 1 8 θ and - 3.2‰ in δ1 3C forpoorly preserved Inoceramus specimens from Upper Cre-taceous strata in Hole 53OA in the Angola Basin of theSouth Atlantic. In the study by Saltzman and Barron(1982), the isotopic values obtained from well-preservedLate Cretaceous specimens ranged from -2.41 to +1.88for δ 1 8 θ and - 1.27 to +3.01 for δ1 3C.
The 90% rejection rate of Inoceramus specimens inthe Saltzman and Barron (1982) study and the observedvariability in the oxygen isotopic composition of the Ino-ceramus specimens listed in Table 3 encourage cautionin the interpretation of paleotemperatures derived fromoxygen isotopic compositions. Some of the Inoceramussamples may have diagenetic alterations that were not
1013

J. A. HAGGERTY
; : •• &
• * * * * - t
' • ' * ' *
Figure 12. A. Scattered skeletal calcite prisms in laminated limestone (Sample 603B-78-3, 65-66 cm) (reflected light; bar scale = 0.5 mm). B. Bor-ings by infaunal organism into macro fossil shell in sediment (Sample 6O3B-78-3, 65-66 cm; lower bedding plane) (reflected light; bar scale =0.5 mm). C. Infilling of borings on upper bedding plane surface from the sample viewed in Figure 12B (Sample 603B-78-3, 65-66 cm) (reflectedlight; bar scale = 0.5 mm). D and E. Mirror images of the upper and lower bedding plane surface showing evidence of boring and bioturbationby infaunal organisms. (Sample 603B-78-3, 65-66 cm) (reflected light; bar scale = 1 mm).
detected by microscope evaluation of the hand specimen.This author recommends against the use of data fromspecimens that have oxygen isotopic compositions morenegative than the host rock value and that also have rel-atively depleted carbon isotopic signatures.
The reason for discounting oxygen isotopic composi-tions that have values more negative than the host rock
value is that such negative values suggest a diagenetic re-equilibration has occurred. Diagenesis may have takenplace later in the burial history of the sediment associ-ated with warmer temperatures or the isotopic composi-tion of the shells and sediment may also be altered bywaters depleted in 18O from clay mineral diagenesis orbasalt alteration. Diagenesis during burial or interaction
1014

PETROLOGY AND CARBON AND OXYGEN STABLE ISOTOPIC COMPOSITION
Table 3. Stable isotope analyses of macro fossils and host rock fromUnit V of Hole 603B.
Core-Section,
interval orlevel in cm
50-1, 124-12653-1, 51-55J>61-1, 86-91°62-2, 50-56b
62-2, 56-57b
63-2, 61-65
76-1, 120
77-1, 48-5278-1, 51-5278-3, 65-6679-1, 10979-1, 11079-2, 58-5979-6, 56-5780-1, 6-780-3, 20-21
81-5, 15-17
Type of rockor macrofossila
InoceramusLaminated limestoneBioturbated limestoneBioturbated limestoneBioturbated limestoneInoceramus
Base of Subunit VA
BelemniteInoceramusInoceramusAptychusBioturbated limestoneAptychusInoceramusInoceramusInoceramusInoceramusInoceramus
Host rock
δ 1 3 C
-0.4-0.3-0.1+ 0.2+ 0.2
+ 0.8+ 1.2+ 0.8+ 0.8-0.7+ 0.9-0.4+ 1.0+ 0.3
-0.4
δ 1 8 θ
-2.7-3.7-4.4-4.5-4.2
-4.1-3.0-2.2-1.9-3.9-2.3-2.7-2.4-2.3
-3.5
Macrofossil
δ 1 3 C
+ 0.4
+ 0*5
+ 0.4- 0 . 3 c
-2.1-0.1
-1.2+ 0.9-0.7-0 .2 c
+ 1.7+ 0.9
δ 1 8 θ
- 2 . 1 d
-4.0
+ 0.2d
- 3 . 3 C
-2.5-7.8
~4JA- 2 . 4 d
-3.2
~ 3 5Λ- 0 . 7 d
- 2 . 7 d
a See Table 1 for the type of host lithology containing the macrofossil.Data from Prezbindowski and Pittman (this volume).
c Hand samples that show distinct evidence of recrystallization.Reliable samples for paleotemperature determination.
with fresh water will deplete the 18O content of the car-bonate. The host rock at Site 603 is diagenetically al-tered (Prezbindowski and Pittman, this volume), andtherefore it is possible that the macrofossils have alsobeen altered.
After the criterion for the oxygen isotopic composi-tion is met, the sample should be evaluated by a secondcriterion based on the carbon isotopic composition. De-pletion of 13C in carbonates commonly results from theincorporation of isotopically light carbon derived fromthe diagenesis of organic matter. Usually the more nega-tive the δ13C composition of the sediment, the greaterthe contribution from organic matter.
Use of the above two guidelines eliminates the twosamples of Inoceramus that visually showed evidence ofrecrystallization for use in paleotemperature determina-tion. Of the remaining seven specimens of Inoceramusthat were analyzed, three of these specimens are alsoeliminated by the above criteria. This may be considereda conservative approach, but most workers (e.g., Ste-vens and Clayton, 1971) have assumed that the lowesttemperature calculated for a sample, locality, or timespan yields the lowest probability of having undergonediagenetic alteration and therefore is closest to the cor-rect value. The specimens that are eliminated for paleo-temperature consideration will typically yield higher ox-ygen isotope paleotemperatures than the remaining "re-liable" specimens.
The oxygen stable isotopic composition of the apty-chi are very depleted in 18O. According to Lehmann(1979), well-preserved aptychi have a thin, dark layer ofhorny or chitinous material covering the innermost cal-cite layer. Many of the aptychi in these cores exhibited adark layer coating the surface (Fig. 3B). The specimenschosen for isotopic analysis had a dark coating that was
+ 2.0
+4.0
Figure 13. Cross plot of δ 1 3C and δ 1 8 θ analyses listed in Table 3. The symbols indicate the lithology of the hostrock samples, as well as the type of macrofossil fragment analyzed: B = bioturbated limestone, L = lami-nated limestone, R .= visibly recrystallized Inoceramus, I = Inoceramus, A = aptychi, and M = belemnite.
1015

J. A. HAGGERTY
not pyritized (Fig. 4B). The host sediment has under-gone diagenesis, as exhibited by replacement of the radi-olarian tests with ferroan calcite, and partial dissolutionof the coccoliths (Prezbindowski and Pittman, this vol-ume), but the depletion of 18O in the ammonite aptychimay not be completely the result of diagenetic re-equili-bration within the sediment.
If one compares the isotopic values of the host rockwith those of the aptychi, the isotopic values of the ap-tychi are significantly depleted. If the aptychi had dia-genetically re-equilibrated with the host sediment, onewould expect the oxygen isotope values to shift towardthe host rock values. The oxygen isotopic compositionof the aptychi may indicate a diagenetic re-equilibrationat a later, warmer time in the burial history of the sedi-ment. Another possible explanation that should be con-sidered is that the initial oxygen isotopic composition ofthe aptychi may reflect a "vital effect" fractionation bythe living organism. If this is the case, the isotopic com-position of the aptychi was never in isotopic equilibriumwith Cretaceous seawater.
The carbon isotopic signature provides another cluethat the aptychi may have undergone some diageneticalteration or are exhibiting additional complications frombiologic fractionation of the isotopes. Both of the speci-mens have negative carbon isotopic signatures. Altera-tion or a slight deterioration of the chitinous organiclayer that was originally in contact with the aptychi mayhave contributed to the negative carbon isotopic signa-ture of the calcite aptychi. A similar depleted carbonisotopic signature is seen in the visibly recrystallized sam-ples of Inoceramus. The two specimens have a negativecarbon isotopic signature, whereas most of the Inocera-mus specimens that were not suspected of being diagenet-ically altered have a positive carbon isotopic signature.
Paleotemperature Results
The Inoceramus specimens exhibit a wide range ofvariability in their isotopic composition. The specimensfrom Subunit VB have an average value of - 1.3%o forδ 1 8 θ and the host rock has an average value of -2.6%ofor δ 1 8 θ. The average value of -1.7‰ for δ 1 8 θ is ob-tained for all the Inoceramus specimens analyzed in UnitV (excluding the two specimens noted earlier as havingbeen recrystallized), whereas the laminated limestone hostrock of Unit V has an average value of *- 2.8%o for δ 1 8 θ .The oxygen isotopic composition of the Inoceramus doesnot appear to be in equilibrium with the host rock.
The isotopic composition of four of the seven Inoce-ramus specimens listed in Table 3 (the specimens thatvisually showed no sign of diagenetic alteration) is con-sistent with the two evaluating criteria discussed in theDiagenetic Trends section. The calculated oxygen iso-tope paleotemperatures of three of these samples indi-cate warm bottom-water temperatures of about 21 to23°C, the fourth gives a value of about 15°C.
Sample 603B-80-3, 20-21 cm, which was not visuallysuspected of being recrystallized, has the strongest posi-tive carbon isotopic signature of all the specimens, andan oxygen isotope composition of -O.7%o, which is en-riched in 18O relative to the host rock. This oxygen iso-
tope value yields a warm, but more reasonable, bottom-water temperature of about 15°C. The assumption thatthe lowest temperature calculated for a sample, locality,or time span yields the lowest probability of having un-dergone diagenetic alteration and therefore is closest tothe correct value may indicate that this is the "most reli-able" Inoceramus specimen and oxygen isotope paleo-temperature. This calculated temperature is in good agree-ment with the belemnite sample (Sample 603B-77-1, 48-52 cm), which yields an oxygen isotope paleotempera-ture of approximately 11°C.
The belemnite shell has not undergone recrystalliza-tion of the calcite rostrum. This was verified by obser-vation of the thin section under crossed nicols on a pet-rographic microscrope (Fig. 4A). The coarse texture ofsparry calcite is observed as an infilling within the ros-trum, but no sparry calcite is observed within the crosssection of the rostrum. The pseudoisogyres that extendthrough the rostrum are also visible under crossed nic-ols. This feature is additional evidence that the micro-structure is not disturbed and that this belemnite is notrecrystallized. The oxygen isotopic composition of thebelemnite is probably indicative of the initial composi-tion of the shell. The author interprets the oxygen iso-tope paleotemperature derived from the isotopic valueof the belemnite to be an appropriate indicator of thetemperature of the water in the Early Cretaceous NorthAtlantic Ocean.
If the carbon isotopic compositions of the four "reli-able" Inoceramus samples are evaluated, those containedin. Subunit VB are enriched in 13C (mean = +1.2), com-pared to those in Subunit VA ( + 0.4). Many times suchsignals, which may be interpreted as climatic, cannot beunraveled from the sediment because of variable com-positions of the sediment components contributing tothe isotopic signature. The Inoceramus should respondas a biologic indicator because the shell should have in-corporated the isotopic signature from the water column.The small number of samples does not permit a distinc-tion as to whether this shift is significant or even reli-able. From the oxygen isotopic composition of the four"reliable" Inoceramus fragments there does not appearto be any distinct trend through time. The question ofchanges in the bottom-water temperature from the lateBerriasian to the Aptian cannot be addressed with suchfew samples. These questions could be answered if a suf-ficient number of unaltered Inoceramus samples wererun and compared with the host sediment. From the da-ta listed in Table 3, there does not appear to be any defi-nite trends. Additional analyses need to be made to con-sider these points.
DISCUSSION
The increase in clay content in the laminated lime-stone compared to the clay content in the bioturbatedlimestone is interpreted as the result of an increase interrigenous sediment input. The laminated limestone con-tains marine organic material as well as abundant terres-trial organic material (Organic Geochemistry section inSite 603 chapter; Dunham et al., this volume). The peri-odic influx of clay, very fine grained sand, and terrige-
1016

PETROLOGY AND CARBON AND OXYGEN STABLE ISOTOPIC COMPOSITION
nous organic matter is interpreted to have resulted in theinterbedded nature of the laminated limestone with thebioturbated limestone at Site 603.
Careful review of the percent of the core containingintervals of laminated limestone versus the other litho-logic types reveals an increase in the laminated limestoneor marl from the bottom of Unit V to the base of Sub-unit VA where the claystone turbidites appear. These in-tervals of laminated limestone anteceded the claystoneturbidites and perhaps indicate the progradation of theTithonian-Barremian deltas (Jansa et al., 1979) that de-veloped on the inner and middle continental shelf. Notuntil the deposition of Subunit VB had the deltas evolvedto the extent that an associated submarine fan could reachthe location of Site 603 on the basin plain. (See Sartiand von Rad, this volume, for a detailed discussion ofthe submarine fan facies.)
This periodic influx of terrestrially derived sedimentassociated with the laminated limestone at DSDP Site603 may have initially been transported from prodeltadeposits by landward-generated sediment gravity flows.Coarser elastics and the majority of the transported sed-iment from these currents would have been deposited onthe shelf or would have aided in the seaward growth ofthe continental slope. Studies of modern sedimentaryprocesses have shown that high-velocity bottom currentscan stir up the seafloor periodically and are capable oftransporting sands for distances before redeposition,whereas finer particles are injected into the nepheloidlayer (Biscaye and Eittreim, 1977; Kennett, 1982). Thissuspension of lutite, if in sufficient quantity, can inducedownslope flow (Ewing and Thorndike, 1965), produc-ing a very weak current. The sediment carried in thenepheloid layer may be transported over greater distancesof the basin plain area and then slowly settle out of sus-pension. Transport of sediment within the nepheloid lay-er may explain the origin of the interbedded laminatedlimestone with the bioturbated limestone at Site 603.
If the laminated limestone in Subunit VB is indeedassociated with the development of the Tithonian-Bar-remian deltas on the continental shelf, then supportingevidence should also appear in the sedirnents as we pro-gress uphole. The clay mineralogy of the laminated lime-stone should progressively change uphole, because thesite would be closer to the source as the deltas prograd-ed seaward. As pointed out by Holmes (this volume),sorting of the clay minerals by mineral species occursbefore deposition. On the basis of size and shape, ka-olinite would settle out closer to the source area, where-as illite would be transported farther away, and smectitewould be transported the farthest from the source area.No clay mineralogy analysis of the laminated limestonewas conducted for this chapter. Selected samples of Cre-taceous strata from Hole 603B were analyzed for claymineralogy by Huff (this volume). Presented in Table 4are data from just the laminated limestones of Unit V atSite 603 from Huff (this volume). These few data sug-gest a minor change in the clay mineral content: a de-crease in smectite, with an associated increase in illite,and a gradual appearance of kaolinite as one progress-es uphole. To prove that a trend of increasing kaolinite
Table 4. Clay mineralogy of laminated limestones fromHole 603B data from Huff (this volume).
Core-Section, Sub-bottomPercent of <2-µm
clay fraction
level in cm
52-1, 123-12652-2, 21-2354-2, 96-10066-5, 70-7576-1, 77-80
(m)
1292.41292.91312.91430.41512.3
Kaolinite
216320
Illite
620212124
Smectite
9264767776
76-1, 120 Base of Subunit VA
78-4, 2-680-2, 35-40
1535.21551.1
8690
content occurs from the base of Subunit VB throughSubunit VA would require numerous analyses of the lime-stone. The bioturbated limestone, laminated limestone,and transitional limestone also display a trend of increas-ing organic carbon content from Subunit VB throughSubunit VA. These data could also be interpreted as theresult of the progradation of the delta over the continen-tal shelf.
The periodic influxes of terrigenous sediments wouldhave increased the contribution of nutrients and organicmatter available as food for benthic organisms. Burrowsthat are infilled with clay-rich sediment are common inthe bioturbated limestone at Site 603. The clay-rich sedi-ment probably was obtained by infaunal organisms fromthe overlying surface sediment, which was enriched interrestrially derived material or was later filled passivelywith this overlying sediment. The bioturbated limestoneat Site 603 always has low organic carbon content be-cause any available organic detritus was probably con-sumed by the infauna.
As the accumulation of the organic material increasedwith the terrestrially derived sediment, the dissolved ox-ygen content of the interstitial waters would have be-come depleted as the result of oxidation of organic com-pounds. The interstitial waters may have become anaer-obic, but some burrowers could have survived if theymaintained a free connection with oxygenated bottomwaters (Ekdale et al., 1984). As the organic detritus con-tinued to be deposited, the sediment/water interface mayhave become a dysaerobic environment with the contin-ued depletion of available dissolved oxygen; a chemical-ly reducing environment would then have been gener-ated within the sediments. Laminated strata that con-tain no macrobenthic fauna are usually interpreted asanaerobic facies because both epibenthic organisms, andinfaunal organisms that would have destroyed the lami-nations, require oxygenated bottom waters (Savrda andBottjer, 1986).
Rhoads and Morse (1971) studied the distribution anddiversity of benthic organisms in modern oxygen-defi-cient basins and compared the results with trace-fossilassemblages from pre-Cambrian and Cambrian strata.From their study, they suggested that as the dissolvedoxygen content of the bottom water decreases, the sizeof the infaunal organisms should also decrease. This hy-pothesis has been tested by Savrda et al. (1984), and this
1017

J. A. HAGGERTY
trend of decreasing burrow size related to dissolved oxy-gen content has been recognized with Chondrites in thePhanerozoic rock record (Bromley and Ekdale, 1984).In addition to the decrease in burrow size with decreas-ing dissolved oxygen content, Savrda and Bottjer (1986)have established and modelled the tiering relationshipsof the infauna with variations in the paleo-oxygenationin bottom waters.
At DSDP Site 603, the ichnofabric of the bioturbatedlimestone does show a distinct decrease in burrow sizewhen approaching the overlying laminated limestone,and a tiering relationship of Planolites crosscut by Chon-drites. Both of these observations fit with the model ofSavrda and Bottjer (1986) for decreasing oxygen contentof the bottom waters. The existence of very small bur-rows of Chondrites within the laminated limestone atDSDP Site 603 is also evidence of very low oxygen levelsin the interstitial waters. Chondrites is capable of exist-ing in sediment characterized by an oxygen content be-low the levels required for other benthic organisms andusually represents the last trace fossil in a tiering rela-tionship (Bromley and Ekdale, 1984; Savrda and Bott-jer, 1986).
The decline in the dissolved oxygen content of thebottom waters associated with the onset of depositionof the laminated limestone at Site 603 is documented bythe decrease in the burrow size and ichnofaunal tieringrelationship in the bioturbated limestone. The dysaero-bic environment of the laminated limestone generatedfrom the continued depletion of available dissolved oxy-gen produced a chemically reducing environment withinthe sediment. The existence of this reducing environmentwithin the laminated strata is supported by the variationin the pyrite content of the sediments. X-ray diffractiondata (Mark Johns, personal communication, 1986) showan increase in the pyrite concentration in the laminatedlimestone by comparison to the bioturbated limestone.In addition, some specimens of Inoceramus were parti-ally pyritized after deposition in the laminated limestone.
The association of Inoceramus with the laminated lime-stone also supports the interpretation that this is not ananaerobic facies. Thiede and Dinkelman's (1977) re-eval-uation of the ecology of Inoceramus shows that theseorganisms were epibenthos that lived in a wide range ofpaleodepths and associated with a variety of sediments,preferring soft mud bottoms. Review of the occurrencesof Inoceramus in DSDP cores reveals that the majorityof the occurrences are associated with laminated, darkmudstones and chalks containing a scarcity of bioturba-tion (Thiede and Dinkelman, 1977) and are therefore in-terpreted to have been tolerant of dysaerobic environ-ments.
The macrofossils or Inoceramus shells at Site 603 arenot interpreted to have been transported downslope tothe basin by the very weak bottom currents associatedwith nepheloid layer that deposited the laminated lime-stone. Thiede and Dinkelman (1977) conservatively lim-ited the paleodepth of Inoceramus to upper bathyal andneritic environments associated with continental and is-land slopes and shelves. They noted that at many DSDPsites the water depth at the time of deposition of Inoce-
ramus is unknown, because the subsidence profile hasnot been determined in many areas. Barron et al. (1984)interpret the paleodepth at DSDP Site 530 to have beenbetween 3500 and 4500 m for the in situ deposition ofInoceramus. Therefore the interpretation of Inoceramusat DSDP Site 603 as being sampled in place and associ-ated with a nearby continental setting does not disagreewith what is known about the ecology of these orga-nisms.
The oxygen isotopic composition of the Inoceramusshells yields very warm, bottom-water temperatures. Thereare two prevalent ideas about bottom-water circulationduring the Cretaceous. One is that the water was slug-gish or stagnant, having been generated in relatively warmpolar regions, and reflecting warm polar surface watertemperatures (Degens and Stoffers, 1976). The other isthat evaporative processes in the subtropics created verywarm dense saline water that dropped to the abyssal floor(Berger, 1979).
The oxygen isotopic composition of biogenic and non-biogenic carbonate precipitated in isotopic equilibriumwith seawater is interpreted to reflect changes in seawa-ter paleotemperature with the assumption that there hasbeen no local variations in salinity. A difference in salin-ity of l%o implies a difference of approximately 1°C forthe calculated paleotemperature (Craig, 1966). Evapora-tion tends to enrich residual water with the heavier oxy-gen isotope, 18O. Calculated paleotemperatures of shellsprecipitated in water that was more saline than thoughtwill be less than the actual temperature (Mook, 1971;Saltzman and Barron, 1982).
The lowest oxygen isotope temperature calculated froma "reliable" Inoceramus shell is 15°C. The temperaturecalculated from the belemnite, which was a nektonic or-ganism, yielded a slightly cooler value of 11°C. All thecalculated temperatures are very warm. Either all of thesamples are isotopically altered or they were not precipi-tated in isotopic equilibrium, or the bottom waters wereindeed very warm.
Saltzman and Barron (1982) obtained temperaturesfrom Inoceramus at Site 530 in the Angola Basin thatthey interpreted as evidence of saline bottom water gen-erated by evaporation. They calculated oxygen isotopepaleotemperatures on the order of 5 to 8°C for Pacificand Indian ocean sites with Upper Cretaceous strata con-taining Inoceramus and interpreted the bottom water inthose areas as having been generated in the polar re-gions. From the temperatures calculated in this study, itappears that they fit more closely with bottom water gen-erated in a warm equatorial region. This bottom water,although not necessarily anoxic, would contain less oxy-gen than polar-generated bottom water. The associationof Inoceramus with only laminated sediment at Site 603may indicate its preference to thrive in bottom waters ofreduced oxygen content (Theide and Dinkelman, 1977).This preferred ecologic niche may explain the absence ofthese organisms in the bioturbated (well-oxygenated) nan-nofossil limestone.
Robertson and Bliefnick (1983) and Robertson (1984)evaluated several depositional scenarios to explain thecyclicity of the laminated limestones with the bioturbat-
1018

PETROLOGY AND CARBON AND OXYGEN STABLE ISOTOPIC COMPOSITION
ed limestones that appeared at DSDP Site 534 in theBlake-Bahama Formation. The postulated causes rangedfrom diagenetic segregation, to the influence of distalturbidites, to current effects, to variations in surface pro-ductivity and influx of clay. None of these theories at-tempted to reconcile the paleoecology of the benthic or-ganisms.
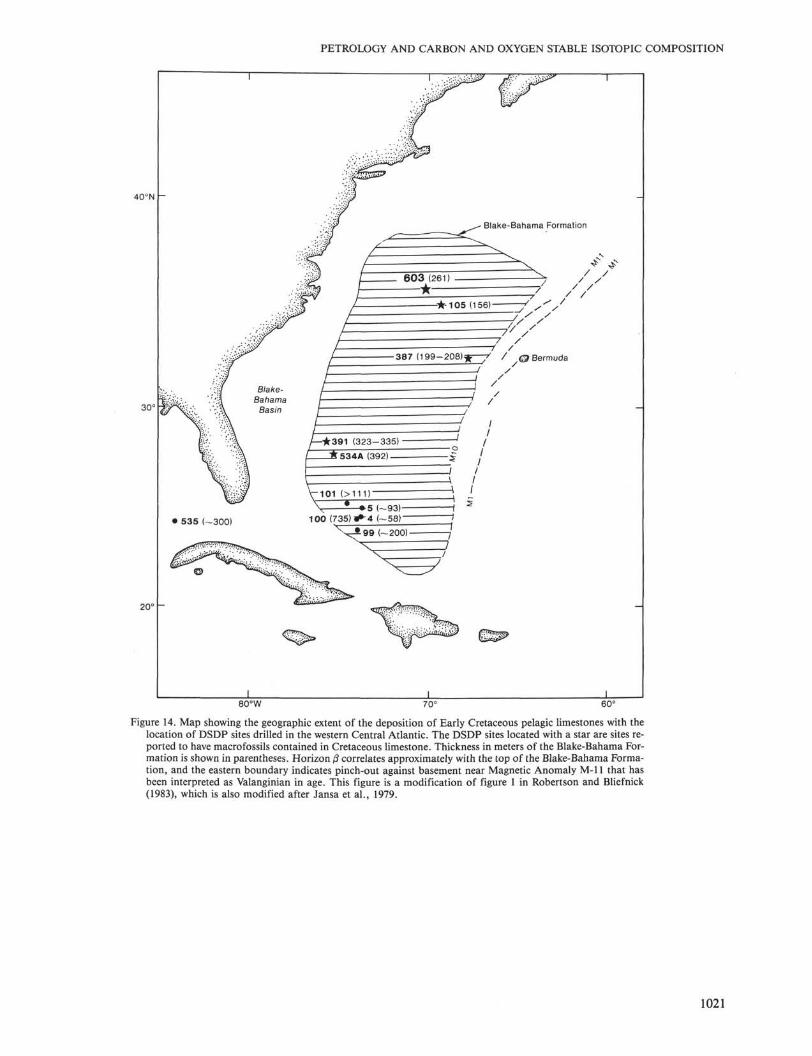
The association of macrofossils with laminated lime-stones from the Blake-Bahama Formation occurs in oth-er locations in the North Atlantic (Fig. 14; Renz, 1972,1978, 1979, 1983; Benson, Sheridan, et al., 1978; Hol-lister, Ewing, et al., 1972; Sheridan, Gradstein, et al.,1983; Tucholke, Vogt, et al., 1979). All of these siteshave the sediment with the relatively largest abundanceof macrofossils (that are predominantly Inoceramus) oc-curring before the onset of deposition of the black clay-stones. At DSDP Site 534, the lithology changes down-hole through the most complete and thickest recoveredsection of the Blake-Bahama Formation, with the disap-pearance of the black claystones followed by the disap-pearance of the parallel-laminated limestones above thebase of the unit (Sheridan, Gradstein, et al., 1983). Allof the drill sites that recovered the Blake-Bahama For-mation in the North Atlantic (Fig. 14) have laminatedlimestones appearing before the deposition of the clay-stone turbidites. Perhaps all of the laminated limestonesof the Blake-Bahama Formation were deposited by veryweak bottom currents associated with the nepheloid lay-er and may have been distantly related to the develop-ment of the Tithonian-Barremian deltas on the innerand middle continental shelf.
CONCLUSIONS
The periodic influx of clay, very fine grained sand, andterrigenous organic matter is interpreted to have resultedin the interbedded nature of the laminated limestone withthe bioturbated limestone at Site 603. The deposition ofintervals of laminated limestone associated with biotur-bated limestone anteceded the deposition of claystoneturbidites and perhaps indicates the progradation of theTithonian-Barremian deltas that developed on the innerand middle continental shelf. Not until the depositionof Subunit VB had the deltas sufficiently developed tohave an associated submarine fan reach the location ofSite 603 on the basin plain. The periodic influx of ter-restrially derived sediment associated with the laminatedlimestone at DSDP Site 603 may have initially been trans-ported from prodelta deposits and subsequently beentransported within the nepheloid layer. Deposition of theseinfluxes of terrigenous detritus from the nepheloid lay-er may explain the origin of the interbedded laminatedlimestone and bioturbated limestone at Site 603. The Ino-ceramus shells at Site 603 are interpreted to be in situ;these have not been transported downslope to the basinby the very weak bottom currents associated with thenepheloid layer that deposited the laminated limestone.
The decline in the dissolved oxygen content of thebottom waters associated with the onset of depositionof the laminated limestone at Site 603 is documented bythe decrease in the burrow size and ichnofaunal tieringrelationship in the bioturbated limestone. The associa-
tion of Inoceramus with the laminated limestone alsosupports the interpretation that this is a dysaerobic fa-cies.
The lowest oxygen isotope temperature calculated froma "reliable" Inoceramus shell is 15°C. The temperaturecalculated from the belemnite, which was a nektonic or-ganism, yielded a slightly cooler value of 11°C. All thecalculated oxygen isotope paleotemperatures are verywarm. Either all of the samples are isotopically altered,or they were not precipitated in isotopic equilibrium, orthe bottom waters were indeed very warm. If these pa-leotemperatures calculated from the Inoceramus are re-liable, then the bottom waters were very warm and mayhave been generated in the equatorial region.
ACKNOWLEDGMENTS
The author thanks her shipboard colleagues for stimulating andinteresting discussions about the Blake-Bahama Formation. Stable iso-tope compositions were determined at the Stable Isotope Laboratoryat the AMOCO Production Company, Research Center. Organic geo-chemistry data were provided by Phil Meyers, and X-ray diffractiondata were provided by Mark Johns. This paper benefited from the re-views by Dennis Prezbindowski, Paul Aharon, and an anonymous re-viewer. The author also acknowledges the University of Tulsa for re-ceipt of a grant from the Faculty Reseach Program to support thisstudy.
REFERENCES
Barron, E. H., Saltzman, E. S., and Price, D. A., 1984. Occurrenceof Inoceramus in the South Atlantic and oxygen isotopic paleo-temperatures in Hole 53OA. In Hay, W. W., Sibuet, J . -C, et al.,Hole 530A. Init. Repts DSDP, 75, Pt. 2: Washington (U.S. Govt.Printing Office), 893-904.
Benson, W. E., Sheridan, R. E., et al., 1978. Init. Repts. DSDP, 44:Washington (U.S. Govt. Printing Office).
Berger, W. H., 1979. Impact of deep sea drilling on paleoceanography.In Talwani, M., Hay, W., and Ryan, W. B. F. (Eds.), Deep DrillingResults in the Atlantic Ocean: Continental Margins and Paleoenvi-ronment: Washington, D.C. (Am. Geophys. Union), Maurice Ew-ing Series, 3:297-314.
Biscaye, P. E., and Eittreim, S. L., 1977. Suspended paniculate loadsand transports in the nepheloid layer of the abyssal Atlantic Ocean.Mar. Geol., 23:155-172.
Bromley, R. G., and Ekdale, A. A., 1984. Chondrites: a trace fossil in-dicator of anoxia in sediments. Science, 224:872-874.
Craig, H., 1966. Isotopic composition and origin of the Red Sea andSalton Sea geothermal brines. Science, 154:1544-1548.
Degens, E. T., and Stoffers, P., 1976. Stratified waters as a key to thepast. Nature, 263:22-27.
Eckdale, A. A., Bromley, R. G., and Pemberton, S. G., 1984. Ichnol-ogy: The Use of Trace Fossils in Sedimentology and Stratigraphy:Tulsa (Soc. Econ. Paleontol. Mineral.), SEPM Short Course No.15.
Ewing, M., and Thorndike, E. M., 1965. Suspended matter in deepocean water. Science, 147:1291-1294.
Gieskes, J.M., 1981. Deep-sea drilling interstitial water studies: impli-cations for chemical alteration of the oceanic crust, Layers I andII. In Warmes, J. E., Douglas, R. G., and Winterer, E. L. (Eds.),The Deep-Sea Drilling Project: A Decade of Progress. Soc. Econ.Paleontol. Mineral. Spec. Publ., 32:149-167.
Hollister, C. D., Ewing, J. I., et al., 1972. Init. Repts. DSDP, 11:Washington (U.S. Govt. Printing Office).
Jansa, L. F., Enos, P., Tucholke, B. E., Gradstein, F. M., and Sheri-dan, R. E., 1979. Mesozoic-Cenozoic sedimentary formations ofthe North American Basin; western North Atlantic. In Talwani,M., Hay, W., and Ryan, W. B. F. (Eds.), Deep Drilling Results inthe Atlantic Ocean: Continental Margins and Paleoenvironment:Washington, D.C. (Am. Geophys. Union), Maurice Ewing Series,3:1-57.
Kennett, J. P., 1982. Marine Geology: Englewood Cliffs (Prentice-Hall).
1019

J. A. HAGGERTY
Lehmann, U , 1979. Aptychus, Anaptychus. In Fairbridge, R. W., andJablonski, D. (Eds.), Encyclopedia of Paleontology: Stroudsburg,PA (Dowden, Hutchinson, and Ross, Inc.), Encyclopedia of EarthSciences, 7:45-46.
Lowenstam, H., and Epstein, S., 1954. Paleotemperatures of the post-Aptian Cretaceous as determined by oxygen isotope method. J.Geoi, 62:207-248.
McCrea, J. M., 1950. On the isotopic chemistry of carbonates and apaleotemperature scale. J. Chem. Physics, 18:849-857.
Majewske, O., 1974. Recognition of Invertebrate Fossil Fragments inRocks and Thin Sections: Netherlands (E. J. Brill), InternationalSedimentary Petrographical Series, 8.
Mook, W. G., 1971. Paleotemperatures and chlorinities from stablecarbon and oxygen isotopes in shell carbonate. Palaeogeogr. Pa-laeoclimatol., Palaeoecol., 9:245-263.
Mutti, E., and Ricci Lucchi, F , 1972. Le torbiditi delPAppennino set-tentrionale: introduzione all'analisi di facies. Mem. Soc. Geol. Ital.,pp. 161-199. (Translated into English: 1978. Turbidites of the North-ern Apennines: introduction to facies analysis. Int. Geol. Rev., 20:125-166.)
Renz, O., 1972. Aptychi (Ammonoidea) from the Upper Jurassic andLower Cretaceous of the western North Atlantic (Site 105, Leg 11,DSDP). In Hollister, C. D., Ewing, J. I., et al., Init. Repts. DSDP,11: Washington (U.S. Govt. Printing Office), 607-630.
, 1978. Aptychi (Ammononoidea) from the Early Creta-ceous of the Blake-Bahama Basin, Leg 44, Hole 391C, DSDP. InBenson, W. E., Sheridan, R. E., et al., Init. Repts. DSDP, 44:Washington (U.S. Govt. Printing Office), 899-909.
_, 1979. Aptychi (Ammonoidea) and ammonites from theLower Cretaceous of the Western Bermuda Rise, Leg 43, Site 387,DSDP. In Tucholke, B. E., Vogt, P. R., et al., Init. Repts. DSDP,43: Washington (U.S. Govt. Printing Office), 591-597.
_, 1983. Early Cretaceous Cephalopoda from the Blake-Baha-ma Basin (Deep Sea Drilling Project Leg 76, Hole 534A) and theircorrelation in the Atlantic and southwestern Tethys. In Sheridan,R. E., Gradstein, F. M., et al., Init. Repts. DSDP, 76: Washington(U.S. Govt. Printing Office), 639-644.
Rhoads, D.C., and Morse, J. W., 1971. Evolutionary and ecologic sig-nificance of oxygen-deficient marine basins. Lethaia, 4:413-428.
Robertson, A. H. F., 1984. Origin of varve-type lamination, gradedclaystones and limestone-shale "couplets" in the Lower Cretaceousof the western North Atlantic. In Stow, D. A. V., and Piper, D. J.W. (Eds.), Fine-Grained Sediments: Deep- Water Processes and Fa-cies: Boston (Blackwell Scientific), pp. 437-452.
Robertson, A. H. F., and Bliefnick, D. M., 1983. Sedimentology andorigin of Lower Cretaceous pelagic carbonates and redeposited elas-tics, Blake-Bahama Formation, Deep Sea Drilling Project Site 534,Western Equatorial Atlantic. In Sheridan, R. E., Gradstein, F. M.,et al., Init. Repts. DSDP, 76: Washington (U.S. Govt. Printing Of-fice), 795-827.
Saltzman, E. S., and Barron, E. J., 1982. Deep circulation in the LateCretaceous: oxygen isotope paleotemperatures from Inoceramus re-mains in DSDP cores. Palaeogeogr., Palaeoclimatol., Palaeoecol.,40:167-181.
Savrda, C. E., and Bottjer, D. J., 1986. Trace-fossil model for recon-struction of paleo-oxygenation in bottom waters. Geology, 14:3-6.
Savrda, C. E., Bottjer, D. J., and Gorsline, D. S., 1984. Developmentof a comprehensive oxygen deficient marine biofacies model: evi-dence from Santa Monica, San Pedro, and Santa Barbara basins,California continental borderland. Am. Assoc. Pet. Geol. Bull.,68:1179-1192.
Shackleton, N. J., and Kennett, J. P., 1975. Paleotemperature historyof the Cenozoic and the initiation of Antarctic glaciation: oxygenand carbon isotope analyses in DSDP Sites 277, 279, and 281. InKennett, J. P., Houtz, R. E., et al., Init. Repts. DSDP, 29: Wash-ington (U.S. Govt. Printing Office), 743-755.,
Sheridan, R. E., Gradstein, F. M., et al., 1983. Init. Repts. DSDP, 76:Washington (U.S. Govt. Printing Office).
Stevens, G. R., and Clayton, R. N., 1971. Oxygen isotope studies onJurassic and Cretaceous belemnites from New Zealand and theirbiogeographic significance. N.Z. J. Geol. Geophys., 14:829-897.
Thiede, J., and Dinkelman, M. G., 1977. Occurrence of Inoceramusremains in late Mesozoic pelagic and hemipelagic sediments. InSupko, P. R., Perch-Nielsen, K., et al., Init. Repts. DSDP, 39:Washington, D.C. (U.S. Govt. Printing Office), 899-910.
Tucholke, B.E., Vogt, P.R., et al., 1979. Init. Repts. DSDP, 43: Wash-ington (U.S. Govt. Printing Office).
Uchupi, E., 1971. Bathymetric atlas of the Atlantic, Caribbean andGulf of Mexico. Woods Hole Oceanographic Institution Ref. No.71-72.
Weber, J. N., and Woodhead, P. M. J., 1972. Temperature dependenceof oxygen-18 concentration in reef coral carbonates. J. Geophys.Res., 77:463-473.
Yapp, C. J., 1979. Oxygen and carbon isotopic measurements of landsnail shell carbonate. Geochim. Cosmochim. Ada, 43:629-635.
Date of Initial Receipt: 6 August 1985Date of Acceptance: 4 June 1986
1020
PETROLOGY AND CARBON AND OXYGEN STABLE ISOTOPIC COMPOSITION
40°N
30c
20c
80°W 70° 60°
Figure 14. Map showing the geographic extent of the deposition of Early Cretaceous pelagic limestones with thelocation of DSDP sites drilled in the western Central Atlantic. The DSDP sites located with a star are sites re-ported to have macrofossils contained in Cretaceous limestone. Thickness in meters of the Blake-Bahama For-mation is shown in parentheses. Horizon ß correlates approximately with the top of the Blake-Bahama Forma-tion, and the eastern boundary indicates pinch-out against basement near Magnetic Anomaly M-ll that hasbeen interpreted as Valanginian in age. This figure is a modification of figure 1 in Robertson and Bliefnick(1983), which is also modified after Jansa et al., 1979.
1021