33063-33068,1994 vol. of no. issue 269, chemi~y the pp. … · a role for two dna helicases in the...
TRANSCRIPT

THE JOURNAL OF B r o m r c ~ ~ C H E M I ~ Y 0 1994 by The American Society for Biochemistry and Molecular Biology, Inc.
Vol. 269, No. 52, Issue of December 30, PP. 33063-33068,1994 Printed in U.S.A.
A Role for Two DNA Helicases in the Replication of T4 Bacteriophage DNA*
(Received for publication, May 19, 1994, and in revised form, October 19, 1994)
Jack Barry and Bruce AlbertsS From the Department of Biochemistry and Biophysics, University of California, Sun Francisco, Sun Francisco, California 94143-0448
-~
The T4 bacteriophage gene 41 protein is the highly processive DNA helicase of the T4 primosome, a central part of the protein machinery that moves the T4 DNA replication fork. The T4 gene 59 protein accelerates the loading of 41 protein onto DNA covered with 32 protein (the T4 single strand binding protein), and it makes the 41 protein DNA helicase activity rapidly available to catalyze replication fork movement through a DNA dou- ble helix (Barry, J., and Alberts, B. M. (1994) J. BioZ. Chem. 269,33049-33062).
With the aid of the 59 protein, we show that the T4 primosome (the T4 gene 41 and 61 proteins) can move rapidly through a promoter-bound RNA polymerase molecule that would otherwise stop replication fork movement. A second, very different DNA helicase, the T4 dda protein, provides an alternative pathway for repli- cation past this DNA-bound RNA polymerase (Bedinger, P., Hochstrasser, M., Jongeneel, C. V., and Alberts, B. M. (1983) Cell 34,115-123). Combined with other data, these in vitro experiments allow us to propose a model that explains why either the 59 protein or the dda protein, but not both, are required to begin efficient DNA repli- cation inside the T4 bacteriophage-infected cell.
The T4 bacteriophage DNA replication apparatus has been reconstituted from a set of purified proteins and used to cata- lyze replication fork movement in vitro (Morris et al., 1975; Sinha et al., 1980; Nossal and Alberts, 1983; Alberts, 1985; Nossal, 1994). Every organism confronts a set of similar prob- lems in replicating its DNA. Studies of the functions of each of the different subunits in the T4 multiprotein complex thereby provide important models for understanding the replication apparatus of other organisms, including humans (Alberts, 1987; Tsurimoto and Stillman, 1990; Capson et al., 1991; Jarvis et al., 1991; Nossal, 1992; O’Donnell et al., 1992; Young et al., 1992; Hacker and Alberts, 1994a, 1994b). In an accompanying paper (Barry and Alberts, 19941, we have purified the T4 gene 59 protein and discovered that it functions to load the highly processive DNA helicase of the T4 primosome, gene 41 protein, onto single-stranded DNA. The stimulation of 41 protein activ- ity by 59 protein is most dramatic when the DNA is covered with the T4 gene 32 protein. In this communication, we use the 59 protein to reinvestigate a problem that we explored previ- ously with the T4 in vitro system (Bedinger et al., 1983): what happens when a moving replication fork encounters a protein that is tightly bound to the parental DNA template?
* This research was supported by National Institutes of Health Grant GM24020. The costs of publication of this article were defrayed in part by the payment of page charges. This article must therefore be hereby marked “aduertisement” in accordance with 18 U.S.C. Section 1734 solely to indicate this fact.
Fax: 415-476-0806. $ To whom correspondence should be addressed. Tel.: 415-476-4132;
Our recent work has demonstrated that a replication fork can bypass a transcribing RNA polymerase molecule without displacing the growing RNA transcript (Liu et al., 1993). How- ever, our earlier studies had suggested that an additional T4 DNA helicase, the T4 dda protein (Jongeneel et al., 1984a, 1984b; Hacker and Alberts, 19921, was required to prevent the replication fork in our in vitro system from stalling when it encounters an RNA polymerase molecule that is bound to a strong promoter on a DNA template prior to transcription ini- tiation (Bedinger et al., 1983). Similar results were obtained in studies in which the protein encountered was the lac repressor bound to its operator DNA sequence.’ Because the 41 protein did not substitute for the dda protein in this assay, these ex- periments were interpreted as indicating that the 41 protein needs help from a second DNA helicase whenever the replica- tion fork encounters a protein that is bound tightly to the parental DNA (Bedinger et al., 1983).
However, with only the help of the DNA polymerase acces- sory proteins (i.e. in the presence of the T4 44/62 and 45 pro- teins, without 59 protein), the 41 DNA helicase loads onto 32 protein-covered, single-stranded DNA very slowly (Barry and Alberts, 1994). Thus, an alternate explanation for the inability of the 41 protein to move the i n vitro replication fork efficiently past DNA-bound proteins was an ineffective assembly of this DNA helicase onto DNA; in particular, since only a short length of single-stranded DNA had been generated by strand displace- ment DNA synthesis before the block was encountered in Bedinger et al. (1983), there might not have been a long enough “loading zone” of single-stranded DNA for 41 protein engage- ment. In this case, the observed requirement for a second DNA helicase would simply reflect the fact that the dda helicase is a highly distributive enzyme (Jongeneel et al., 1984b) that can be much more readily recruited to the blocked replication forks in our in vitro system than 41 protein.
Our suspicions were raised by the finding that dda- T4 mu- tants synthesize a great deal of DNA in uivo; although these viruses start their DNA synthesis with a short delay, they even- tually produce a normal burst size (Gauss et al., 1994). This observation suggests that a second protein can substitute for the dda protein and move the fork past DNA-bound proteins. When genetic data revealed that viruses deficient in both 59 protein and dda protein make little or no DNA, whereas a virus missing only one of these two protein makes DNA (Gauss et al., 19941, our attention turned to the gene 59 protein as this sec- ond protein.
The 59 protein binds to DNA, to 32 protein, and to 41 protein; in its presence the 41 protein loads rapidly onto DNA (Barry and Alberts, 1994; Morrical et al., 1994; Yonesaki, 1994). In the experiments reported here, we use the 59 protein to show that the 41 protein is able to move the fork past a promoter-bound RNA polymerase molecule. We conclude that, once assembled
’ J. Barry and B. Alberts, unpublished data.
33063

33064 ITzvo Helicases Help Replicate DNA inside the cell, the T4 primosome is sufficient to move a DNA replication fork past DNA-bound proteins. This finding allows us to explain the observation that, in vivo, a 59- mutant has no effect on early DNA synthesis, unless the dda protein is also missing.
EXPERIMENTAL PROCEDURES Reagents, Enzymes, and Nucleic Acids-The T4 bacteriophage dda
DNA helicase, expressed from the cloned gene, was purified as de- scribed (Hacker and Alberts, 1992). Highly purified Escherichia coli RNA polymerase holoenzyme was a generous gift from the laboratory of Michael J . Chamberlin, University of California, Berkeley. All other
Alberts (1994). reagents, enzymes, and the pJMCllO DNA are described in Barry and
Assays for Strand Displacement DNA Synthesis Starting from a Nick on a Circular Double-stranded DNA ILmplate-All reaction mixtures contained assay buffer, deoxyribonucleoside triphosphates (including 0.1 mM [CY-~~PI~TTP at 800 Ci/mol), specifically nicked pJMCllO DNA (serving as both template and primer), and the DNA polymerase ho- loenzyme (T4 bacteriophage 43, 44/62, and 45 proteins) at the concen- trations described previously (Barry and Alberts, 1994). RNA polymer- ase and the T4 bacteriophage 32, 41, and 59 proteins, when present, were used at the concentrations specified in the figure legends. When present, the dda DNA helicase was used at 1.6 pg/ml; the 61 DNA primase was used at 1 pg/ml, but the absence of the four ribonucleoside triphosphates prevented lagging strand synthesis.
In the experiment lacking RNA polymerase (Fig. l), DNA synthesis reactions were carried out at 37 “C as described in Barry and Alberts (1994). Reactions containing RNA polymerase were carried out as de- scribed in Fig. 2. Quantitation of DNA products (incorporation of [w3’P1dTTP into an acid-insoluble form) and measurement of DNA product size by alkaline agarose gel electrophoresis were performed as described in Barry and Alberts (1994).
RESULTS
The genetic evidence suggests that the T4 dda DNA helicase and the T4 gene 59 protein have at least one function in DNA replication that is similar, since mutant viruses that lack both of these gene products are much more defective in their DNA synthesis than are viruses that lack only one (Gauss et al., 1994). Could we obtain a similar result in an in vitro DNA synthesis reaction? The results of such an experiment, carried out in the presence of the gene 41 DNA helicase, are plotted in Fig. 1. As in vivo, the in vitro reaction is stimulated by dda protein (open triangles) nearly as well as by 59 protein (open squares), and little additional stimulation results from the si- multaneous presence of both proteins (solid squares).
Both the 59 Protein and the dda Protein Can Enable a Replication Fork to Pass a Promoter-bound RNA Polym- erase-To further explore the possibility of redundant func- tions, we performed an experiment to address the major ques- tion raised in the Introduction. In our previous T4 in vitro system, the dda DNA helicase was needed to move a replication fork past an RNA polymerase molecule bound to a strong pro- moter, causing this RNA polymerase to be displaced from the DNA (Bedinger et al., 1983). In these early experiments, the 41 DNA helicase, alone or with 61 protein, did not substitute for the dda protein (Bedinger et al., 1983). Would this result be different in the presence of 59 protein? To examine this ques- tion, the original experiment was repeated with and without 59 protein present, as outlined in Fig. 2 A . On the DNA template used, replication starts by the covalent addition of deoxyribo- nucleotides onto the 3’-OH DNA end at a site-specific nick, and it proceeds for about 150 nucleotides before encountering an RNA polymerase molecule bound to promoter A, the strong gene I1 promoter of bacteriophage M13 (Bedinger et al., 1983; Fig. 2 A ) .
The relative amounts of DNA made in this experiment are graphed in Fig. 3. The preincubation of the DNA and the RNA polymerase (17 molecules per DNA template) in a reaction con- taining six of the T4 DNAreplication proteins (32,43,44/62,45,
I
one copy ”“
0 1 2
reaction time (minutes)
FIG. 1. Stimulation of in vitro DNA synthesis by the T4 69 and dda proteins, added either singly or together. Strand displace- ment DNA synthesis was carried out as described under “Experimental Procedures.” All reactions contained 38 pg/ml of 41 protein, 68 pg/ml of 32 protein, and the DNA polymerase holoenzyme (see “Experimental Procedures”). Where indicated, 0.6 pg/ml of 59 protein and 1.6 pg/ml of dda protein were included. DNA synthesis was measured at 1 and 2 min after dCTP addition. “one copy” marks the incorporation expected when one copy of the pJMCll0 DNA template strand (2.2 nmoVml) has been made. For reference, the DNA synthesis with dda protein in the absence of 41 protein was 1.6 and 3.1 nmoVml at 1 and 2 min, respectively (data not plotted). In the absence of both the dda and 41 proteins, DNA
either the presence or absence of 59 protein (data not plotted). Closed synthesis was 0.65 and 1.1 nmoVml at 1 and 2 min, respectively, in
squares, plus 59 and dda proteins; open squares, plus 59 protein; open triangles, plus dda protein; open circles, no addition.
and 41 proteins) shuts down DNA synthesis nearly completely (compare two dashed lines). As in our previously published experiments (Bedinger et al . , 19831, the presence of the 41 DNA helicase was without effect in preventing this shut-down (solid circles), whereas the addition of the dda DNA helicase to the reaction removes much of the inhibition (solid triangles). But an even more striking restoration of DNA synthesis is obtained if the dda protein is omitted, and the 59 protein is added in the presence of 41 protein (open squares in Fig. 3).
When the samples in Fig. 3 were analyzed by alkaline aga- rose gel electrophoresis, so as to measure the DNA chain lengths produced in each reaction, the results shown in Fig. 4 were obtained. Even with 41 protein present, the block in chain elongation at the first promoter encountered by the replication fork (promoter A) is seen to be nearly absolute without 59 protein or dda protein present (compare lanes C and D with lanes A and B ) , although a few forks “leak through” by 2 min (lane D ) . The addition of either the 59 protein (lanes E and F ) or the dda protein (lanes G and H ) causes a dramatic increase in the production of long DNA chains. However, a major differ- ence can be seen: whereas both reactions produce very large DNA products after 2 min of synthesis (compare lanes F and H ) , at 1 min much longer DNA strands are produced in the presence of the 59 and 41 proteins than with the dda and 41 proteins (compare lanes E and GI.
What happens in these same reactions if the 41 protein is omitted? For DNA synthesis blocked by RNA polymerase in the absence of 41 protein, the addition of 59 protein is without effect, leaving all forks blocked as in lanes C and D (in Fig. 4). As observed previously (Bedinger et al., 19831, the addition of the dda DNA helicase allows extensive DNA synthesis. How- ever, the DNApolymerase now moves relatively slowly, produc- ing relatively short DNA strands (Fig. 4, lanes Z and J ) .
By comparing lanes G and H (dda plus 41 protein) with lanes I and J (dda protein alone), we see that 41 DNA helicase is essentially without effect during the first minute of reaction; in both cases the DNA polymerase holoenzyme has extended the nicked primer strand of the 4755 base pair double-stranded
l tvo Helicases Help Replicate DNA 33065
A site-specific nlck & promoter A bound to promoter A
RNA polymerase 5’
pJMCt10 4755 bp
incubate one mln
polymerase RNA polymerase
bound to promoter B
____, add reolication
incubate wlthout proteins+
dCTP lor two min
*;i pomoter A
holoenzyme ardgene 32 dCTP add 4 DNA pol merase
without dCTP for two min synthesis protein; incubate to start
___, add
dGTP to stan
synthesis 0 an RNA polymerase bound replication fork enwunters
to promoter A
an gNA polymerase bound to re ication fork enwunters
promoter A or promoter B
polymerase with or
41 and 61 without
proteins
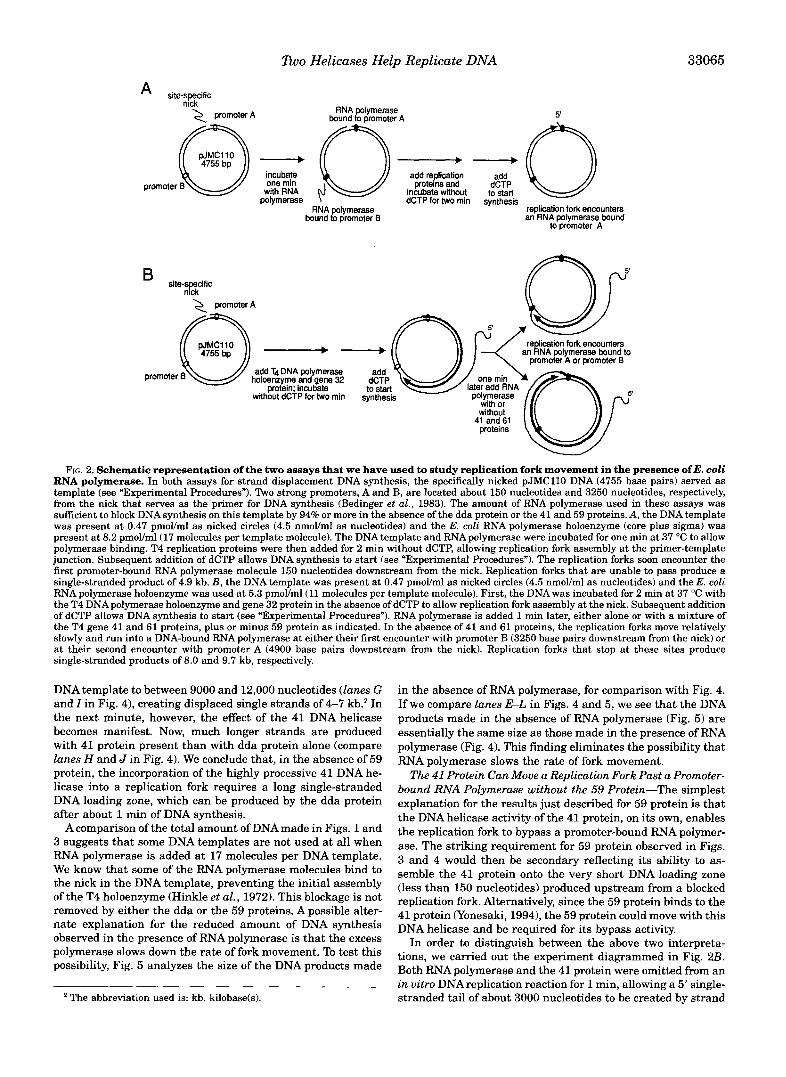
RNA polymerase. In both assays for strand displacement DNA synthesis, the specifically nicked pJMCll0 DNA (4755 base pairs) served as FIG. 2. Schematic representation of the two assays that we have used to study replication fork movement in the presence of E. coli
template (see “Experimental Procedures”). Two strong promoters, A and B, are located about 150 nucleotides and 3250 nucleotides, respectively, from the nick that serves as the primer for DNA synthesis (Bedinger et al., 1983). The amount of RNA polymerase used in these assays was sufficient to block DNA synthesis on this template by 94% or more in the absence of the dda protein or the 41 and 59 proteins. A, the DNA template was present at 0.47 pmoVml as nicked circles (4.5 nmoVml as nucleotides) and the E. coli RNA polymerase holoenzyme (core plus sigma) was present at 8.2 pmoVml(17 molecules per template molecule). The DNA template and RNApolymerase were incubated for one min at 37 “C to allow polymerase binding. T4 replication proteins were then added for 2 min without dCTP, allowing replication fork assembly at the primer-template junction. Subsequent addition of dCTP allows DNA synthesis to start (see “Experimental Procedures”). The replication forks soon encounter the first promoter-bound RNA polymerase molecule 150 nucleotides downstream from the nick. Replication forks that are unable to pass produce a single-stranded product of 4.9 kb. B, the DNA template was present at 0.47 pmoVml as nicked circles (4.5 nmoVml as nucleotides) and the E. coli RNApolymerase holoenzyme was used at 5.3 pmoVml(11 molecules per template molecule). First, the DNA was incubated for 2 min at 37 “C with the T4 DNApolymerase holoenzyme and gene 32 protein in the absence of dCTP to allow replication fork assembly at the nick. Subsequent addition of dCTP allows DNA synthesis to start (see “Experimental Procedures”). RNA polymerase is added 1 min later, either alone or with a mixture of the T4 gene 41 and 61 proteins, plus or minus 59 protein as indicated. In the absence of 41 and 61 proteins, the replication forks move relatively slowly and run into a DNA-bound RNA polymerase at either their first encounter with promoter B (3250 base pairs downstream from the nick) or at their second encounter with promoter A (4900 base pairs downstream from the nick). Replication forks that stop at these sites produce single-stranded products of 8.0 and 9.7 kb, respectively.
DNAtemplate to between 9000 and 12,000 nucleotides (lanes G and Z in Fig. 4), creating displaced single strands of 4-7 kb.2 In the next minute, however, the effect of the 41 DNA helicase becomes manifest. Now, much longer strands are produced with 41 protein present than with dda protein alone (compare lanes H and J in Fig. 4). We conclude that, in the absence of 59 protein, the incorporation of the highly processive 41 DNA he- licase into a replication fork requires a long single-stranded DNA loading zone, which can be produced by the dda protein after about 1 min of DNA synthesis.
A comparison of the total amount of DNA made in Figs. 1 and 3 suggests that some DNA templates are not used at all when RNA polymerase is added at 17 molecules per DNA template. We know that some of the RNA polymerase molecules bind to the nick in the DNA template, preventing the initial assembly of the T4 holoenzyme (Hinkle et al., 1972). This blockage is not removed by either the dda or the 59 proteins. A possible alter- nate explanation for the reduced amount of DNA synthesis observed in the presence of RNA polymerase is that the excess polymerase slows down the rate of fork movement. To test this possibility, Fig. 5 analyzes the size of the DNA products made
The abbreviation used is: kb, kilobaseb).
in the absence of RNA polymerase, for comparison with Fig. 4. If we compare lanes E-L in Figs. 4 and 5, we see that the DNA products made in the absence of RNA polymerase (Fig. 5) are essentially the same size as those made in the presence of RNA polymerase (Fig. 4). This finding eliminates the possibility that RNA polymerase slows the rate of fork movement.
The 41 Protein Can Move a Replication Fork Past a Promoter- bound RNA Polymerase without the 59 Protein-The simplest explanation for the results just described for 59 protein is that the DNA helicase activity of the 41 protein, on its own, enables the replication fork to bypass a promoter-bound RNA polymer- ase. The striking requirement for 59 protein observed in Figs. 3 and 4 would then be secondary reflecting its ability to as- semble the 41 protein onto the very short DNA loading zone (less than 150 nucleotides) produced upstream from a blocked replication fork. Alternatively, since the 59 protein binds to the 41 protein (Yonesaki, 19941, the 59 protein could move with this DNA helicase and be required for its bypass activity.
In order to distinguish between the above two interpreta- tions, we carried out the experiment diagrammed in Fig. 2 B .
Both RNA polymerase and the 41 protein were omitted from an in vitro DNAreplication reaction for 1 min, allowing a 5’ single- stranded tail of about 3000 nucleotides to be created by strand

33066 ltoo Helicases Help Replicate DNA
0 1 2
reaction time (minutes) FIG. 3. In the presence of the T4 gene 41 DNAhelicase, either the
T4 gene 59 protein or the T4 dda DNA helicase can overcome the block to DNA synthesis caused by a promoter-bound RNA po- lymerase molecule. Reactions were carried out as described under“Ex- perimental Procedures” and Fig. 2 A . All reactions contained 68 pgml of 32 protein and the DNApolymerase holoenzyme (see “Experimental Pro- cedures”). The 41 DNA helicase (used at 38 pg/ml) was present in all but one reaction. Where indicated, reactions also contained 0.6 pg/ml of 59 protein, 1.6 pg/ml of dda protein, and 4.1 pg/ml of RNApolymerase. DNA synthesis was measured a t 1 and 2 min after dCTP addition. “one copy” is defined in Fig. 1. Closed squares, plus RNApolymerase and 59 and dda proteins; open squares, plus RNA polymerase and 59 protein; open tri- angles, plus RNA polymerase and dda protein; open circles, no RNA po- lymerase; closed triangles, plus RNA polymerase and dda protein (no 41 protein); closed circles, plus RNA polymerase.
displacement DNA synthesis. If RNApolymerase is then added, i t binds to the two strong promoters present on the plasmid (designated as promoter A and promoter B) within a minute or two, stopping the replication fork a t those sites within 2.5 min (Fig. 6, lane B; see also Fig. 2 B ) . However, if the 41 protein is added along with the RNA polymerase, a race ensues between RNA polymerase binding and 41 protein loading onto DNA. Many replication forks are initially stopped (compare lanes E and H ) . But by 5 min of incubation (4 min after RNA polymer- ase addition), essentially all of the forks have acquired the 41 DNA helicase, bypassed the promoter-bound RNA polymerase molecules, and moved many times around the template DNA circle in the absence of 59 protein (note the loss of the 8.0- and 9.7-kb bands in lane F that were present in lane E of Fig. 6). When 59 protein is added to the reaction along with RNA polymerase and the 41 protein, the replication forks do not stop at the promoters even transiently (compare lanes M and N with lanes D and E in Fig. 6). The RNA polymerase now looses the race from the very start, indicating that the 59 protein accel- erates the incorporation of 41 protein into a replication fork even when thousands of nucleotides of single-stranded DNA are available for this assembly reaction.
As a control for the Fig. 6 experiment, an identical experi- ment was carried out without 41 protein present. The results, presented in Fig. 7, show that 59 protein is without effect if the 41 protein is omitted.
We conclude that the block caused by a promoter-bound RNA polymerase molecule in our in vitro DNA replication system is relieved by the 59 protein only because this protein loads the 41 DNA helicase rapidly onto DNA.
DISCUSSION
In this paper, we have shown that we can use an in vitro DNA synthesis reaction carried out with purified proteins to mimic the observed effects of the T4 59 and dda proteins on in vivo
41 protein + 59 protein -
dda protein - RNA polymerase added -
before DNA synthesis Time (minutes) 1 2
Lane A E
40.0-
23.1 -
DNA size (kb)
9.4 -
+ - - +
1 2 2 D
+ + - +
1 2 E F
+ - + +
1 2 G H
..
- + - + + + + + 1 2 1 2 I J K L ”. .
Block at first 6.6- encounter with promoter A, 4.9 kb
L 4.4 -
FIG. 4. Alkaline agarose gel analysis of the DNA products from the DNA synthesis reactions described in Fig. 3. Samples taken after 1 and 2 min of DNA synthesis were prepared for electrophoresis on a 0.5% agarose gel as described under “Experimental Procedures.” Equal volumes of sample were analyzed in each gel lane; sizes were determined by reference to ”P-labeled marker DNAs. The gel was pre- pared and run as described elsewhere (Formosa and Alberts, 1986). The autoradiograph of the dried gel is shown. RNApolymerase was bound to the DNA prior to the addition of the replication proteins (see Fig. 2A ). Note that [CY-~~PI~TTP was continuously present, so that the large DNA products are disproportionately labeled. Continuous labeling was used in all of the experiments described in this report.
41 protein + 59 protein -
dda protein - Time (minutes) 1 2
Lane A E
40.0-
23.1 - DNA size (kb)
9.4 -
6.6 -
+ + -
1 2 E F
+ - +
1 2 G H
- +
1 2 I J
+ + +
1 2 K L
4.4 - FIG. 5. Alkaline agarose gel analysis of the DNAproducts from
the DNAsynthesis reactions described in Fig. 1. Samples taken a t
0 .58 agarose gel and run as described under “Experimental Proce- 1 and 2 min of DNA synthesis were prepared for electrophoresis on a
dures.” RNA polymerase was not present in these reactions. The samples shown here were fractionated on the same gel as the samples shown in Fig. 4. These DNAproducts are seen to be essentially the same size as those synthesized in the presence of RNA polymerase (compare with Fig. 4). However the average DNA size of the product bands in lanes E-L was about 1 kb larger than the average DNA size in the presence of RNA polymerase (lanes E-L in Fig. 4). indicating that there was a brief pause of 2-3 s before stalled replication forks (paused a t a promoter-bound RNA polymerase) could proceed with the help of 41-59 proteins and/or dda protein. Equal volumes of sample were analyzed in each gel lane. An autoradiograph of the dried gel is shown.

T h o Helicases Help Replicate DNA 33067
of the products from DNA synthesis FIG. 6. Alkaline agarose gel analysis
in an experiment carried out accord- ing to Fig. 2B. In these experiments, the RNA polymerase and 41 protein were added only after 1 min of DNA chain elon- gation. All reactions contained 39 pg/ml of 32 protein and the DNA polymerase ho- loenzyme (see "Experimental Proce- dures"). One min after synthesis had been started by the addition of dCTP, in vari- ous combinations as indicated, 42 pg/ml of 41 protein, 1 pg/ml of 61 protein, and 0.4 pg/ml of 59 protein were added to the re- action, along with 2.5 pg/ml of RNA poly- merase.These proteins had been preincu- bated together with 258 of the reaction's dGTP for 3 min a t 37 "C prior to their addition to the reaction, in order to equili- brate temperature and help activate the 41 protein (Liu and Alberts, 1981). DNA synthesis was stopped a t 2,3.5, and 5 min after dCTP addition. Samples of the reac- tion were prepared for alkaline agarose gel electrophoresis and were fractionated on 0.5% gels as described under "Experi- mental Procedures" and Fig. 4. An auto- radiograph of the dried gel is shown.
41 protein - 59 protein -
RNA polymerase added at one minute -
Time (minutes) 2 3.5 5
Lane A B C
40.0 - 21 .o -
Block at second encounter with 15.0 - promoter A, 9.7 kb
\ 11.0-
Block at first encounter with 7 promoter B, 8.0 kb 7.0 -
6.0 - DNA size (kb) 5.0 -
4.0 -
41 and 61 proteins - 59 protein -
added at one minute i- RNA polymerase
Time (minutes) 2 3.5 5
Lane A B C
40.0 - 21.0 -
Block at second encounter with 15.0 - promoter A. 9.7 kb , .o - Block at first encounter with I-* promoter B, 8.0 kb 7.0 -
6.0 - 5.0 - 4.0 -
L
DNA size (kb)
- - - + + + + -
2 3.5 5
J K L D E F G H I
2 3.5 5 2 3.5 5 FIG. 7. The 59 protein has no effect on replication forks stalled at a pro- moter-bound RNA polymerase in the absence of 41 protein. Alkaline agarose gel analysis of the products from DNA synthesis was camed out as depicted schematically in Fig. 2 B , but with no 41 protein present. Both 41 protein and 61 protein were omitted; otherwise, condi- tions were identical to those described in Fig. 6. Experiments in which 61 protein was omitted show that it is not essential to the 59 protein effect (see, for example, Figs. 3 and 4 and also Barry and Alberts (1994)). Therefore, we conclude that the effect of 59 protein on these stalled repli- cation forks absolutely requires only the 41 DNA helicase.
DNA synthesis. Thus, in vitro DNA synthesis is strongly stimu- lated in the presence of either protein, but the presence of both proteins together provides little added advantage (Fig. 1). This experiment was carried out on a DNA template with an un- natural start site, since replication begins from a nick on a double-stranded DNA circle, rather than from the replication bubble that begins DNA replication in vivo (Mosig, 1983; Kreuzer and Morrical, 1994). Can the Fig. 1 results neverthe- less be used to explain the similar observations made in vivo with T4 mutant-infected cells?
The results in this study allow us to interpret the Fig. 1 result as follows. The loading of the 41 protein onto 32 protein- covered DNA is a rate-limiting step for DNA synthesis, since this DNA helicase not only greatly speeds up the rate of repli-
cation fork movement, but also enables this fork to pass certain proteins such as promoter-bound RNA polymerases that are tightly bound to the DNA. Providing that a nonprocessive DNA helicase, the T4 dda protein, is present, the T4 DNA polymer- ase holoenzyme can proceed far enough without the 41 protein to displace a long region of the template's complementary strand. When this 32 protein-covered DNA single strand at- tains the requisite length (for example, a 3-5-kb extension of the 4.75-kb primer strand after 1 min of synthesis in the ex- periment described in Fig. 2L3 ), the T4 DNA polymerase acces- sory proteins can assemble the 41 protein onto the DNA (Nossal, 1994; Barry and Alberts, 1994). A very rapid DNA synthesis begins as soon as the highly processive 41 DNA he- licase becomes properly engaged with the DNA. This transition

33068 7Iuo Helicases Help Replicate DNA
I I , RNA polymerase replication origin
incompletely characterized initiation proteins
form replication bubble
41 helicase activity promoted by 59 protein RNA polymerase
v
i replication fork
complete
41 heiicase activity promoted by
accessoty proteins
i replication fork
complete
protein, but not both, to obtain T4 DNA synthesis in vivo. As FIG. 8. A model to explain the requirement for either dda or 69
indicated, there are two possible pathways by which the 41 protein becomes incorporated into a replication fork to form the primosome. In the absence of 59 protein, the 41 protein can be loaded less eEciently onto single-stranded DNA by the DNA polymerase accessory proteins (the 44/62 and 45 proteins; see Table I11 in Richardson and Nossal (1989) and Fig. 8 in Barry and Alberts (1994)); this pathway requires a longer loading zone than is needed for loading of the 41 protein by the 59 protein.
to rapid DNA synthesis occurs even more readily if the 59 protein is present, since the 41 protein can be incorporated into a replication fork within seconds (Barry and Alberts, 1994), and even a very short region of displaced single strand (less than 150 nucleotides) suffices as a loading zone (see Figs. 2A and 4).
We have recently shown that the replication fork leaves the RNA polymerase molecule on the DNA when it passes a trun- scribing RNA polymerase (Liu et al., 1993). We have not inves- tigated the fate of the bypassed RNA polymerase examined here, which sits on a strong promoter DNA sequence without a transcript. Although this RNA polymerase was shown to be displaced from the DNA in Bedinger et al. (19831, this result may be attributable to the presence of the dda DNA helicase, rather than to the difference between a promoter-bound and a transcribing RNA polymerase m~lecule.~ Additional experi- ments will be required to decide this issue.
If we apply our in vitro findings to the in vivo situation, we can suggest a model that explains why the 59 and dda proteins can substitute for each other, at least partially, at early times of T4 infection (Gauss et al., 1994). As illustrated in Fig. 8, what
B. Liu and B. Alberts, unpublished data.
ever mechanism is used to prime DNA synthesis at an early T4 replication origin (see Kreuzer and Morrical, 1994), we might expect it to be difficult to advance the leading strand beyond a DNA-bound protein like the promoter-bound RNA polymerase when both 59 and dda proteins are missing. Therefore very little DNA synthesis occurs.
Once a replication bubble forms in a normal infection, the dda protein can clear away DNA-bound proteins, allowing a large single-stranded loading zone to form, onto which the 41 protein eventually assembles with the help of the DNA polym- erase accessory proteins. In the absence of the dda protein, DNA synthesis is somewhat delayed until enough 59 protein accumulates to load the 41 protein on a short region of DNA single strand. In either case, once the 41 protein is incorporated into the replication fork, the fork moves rapidly away from the origin in a highly processive manner, terminating only when all of the DNA ahead of the fork has been replicated.
Acknowledgments-We thank Dr. Scott Morrical, Dr. Robert Schrock, and Bin Liu for valuable discussions about the data and for critical reading of the manuscript. We also thank Fran Dependahl and Rachel Kline for typing the manuscript.
REFERENCES
Alberts, B. M. (1987) Philos. Bans. R. Soc. Lond. B 317, 395-420 Alberts, B. M. (1985) Bends Genet. 1 , 2 6 3 0
Barry, J., and Alberts, B. M. (1994) J. Biol. Chem. 269,3304933062 Bedinger, P., Hochstrasser, M., Jongeneel, C. V., and Alberts, B. M. (1983) Cell 34,
Capson, T. D., Benkovic, S. J., and Nossal, N. G. (1991) Cell 66,249-258 Formosa, T., and Alberts, B. M. (1986) Cell 47, 793-806 Gauss, P., Park, K., Spencer, T. E., and Hacker, K. J. (1994) J. Bacteriol. 176,
Hacker, K. J., and Alberts, B. M. (1992) J. Biol. Chem. 267, 20674-20681 Hacker, K. J., and Alberts, B. M. (1994a) J. Biol. Chem. 269, 24209-24220 Hacker, K. J., and Alberts, B. M. (1994b) J. Biol. Chem. 269,24221-24228 Hinkle, D. C., Ring, J., and Chamberlin, M. J. (1972) J. Mol. Biol. 70, 197-207 Jarvis, T. C., Newport, J. W., and von Hippel, P. H. (1991) J. Biol. Chem. 266,
115-123
1667-1672
111.10-1 1140 Jongeneel, C. V., Bedinger, P., and Alberts, B. M. (1984a) J. Biol. Chem. 269, "" " ~ .
12933-12938 Jongeneel, C. V, Formosa, T., and Alberts, B. M. (1984b) J. Biol. Chem. 269,
12925-12932 Kreuzer, K. N., and Morrical, S. W. (1994) in Molecular Biology of Bacteriophage T4
( h a m , J., ed) pp. 2842 , American Society for Microbiology, Washington, D. C. Liu, C. C., and Alberts, B. M. (1981) J. Biol. Chem. 266,2813-2820 Liu, B., Wong, M. L., Tinker, R. L., Geiduschek, E. P., and Alberts, B. M. (1993)
Morrical. S. W.. Hemustead. IC. and Morrical. M. D. (1994) J. Biol. Chem. 269, Nature 366,33-39
. . . 33069-33081
Moms, C. F., Sinha, N. K., andAlberts, B. M. (1975)Proc. Natl. Acud. Sei. U. S. A.
Mosig, G. (1983) in Bacteriophage T4 (Mathews, C. K., Kutter, E. M., Mosig, G., 72,4800-4804
Washington, D. C. and Berget, P. B., eds) pp. 120-130, American Society for Microbiology,
Nossal, N. G. (1992) FASEB J. 6,871-878 Nossal, N. G. (1994) in Molecular Biology of Bacteriophage T4 ( G r a m , J . , ed) pp.
Nossal, N. G., and Alberts, B. M. (1983) in Bacteriophage T4 (Mathews, C. K., 43-53, American Society for Microbiology, Washington, D. C.
Kutter, E. M., Mosig, G., and Berget, P. B., eds) pp. 71-81, American Society for Microbiology, Washington, D. C.
O'Donnell, M., Kuriyan, J., Kong, X. P., Stukenberg, P. T., and Onrust, R. (1992) Mol. Biol. Cell 3, 953-957
Richardson, R. W., and Nossal, N. G. (1989) J. Biol. Chem. 264,4732-4739 Sinha, N. K., Moms, C. F., andAlberts, B. M. (1980) J. Biol. Chem. 265,4290-4303 Tsurimoto, T., and Stillman, B. (1990) Proc. Natl. Acad. Sei. U. S. A. 87,1023-1027
Young, M. C., Reddy, M. K., and von Hippel, P. H. (1992) Biochemistry 31,8675- Yonesaki, T. (1994) J. Biol. Chem. 269, 1284-1289
8690