25 secondary metabolites: an introduction to natural ... · pdf fileproducts chemistry in the...
TRANSCRIPT

1016Look for this logo in the chapter and go to OrganicChemistryNow
at http://now.brookscole.com/mcmurryorgbio1 for tutorials, simulations, and problems.
25 Secondary Metabolites: An Introduction to NaturalProducts Chemistry
In the past six chapters, we’ve looked at the chemistry and metabolism of thefour major classes of biomolecules—proteins, carbohydrates, lipids, andnucleic acids. But there is far more to do, for all living organisms also containa vast diversity of substances usually grouped under the heading naturalproducts. The term natural product really refers to any naturally occurringsubstance but is generally taken to mean a secondary metabolite—a smallmolecule that is not essential to the growth and development of the produc-ing organism and is not classified by structure.
It has been estimated that well over 300,000 secondary metabolites exist,and it’s thought that their primary function is to increase the likelihood of anorganism’s survival by repelling or attracting other organisms. Alkaloids, suchas morphine; eicosanoids, such as prostaglandin E1; and antibiotics, such aserythromycin and the penicillins, are examples.
Prostaglandin E1
BenzylpenicillinErythromycin A
Morphine
CO2HH
HH OHHOH
H H
OCH3O
H
H
H H
O
HO
HO
HO
O
CH3
CH3
CH3
CO2–
N
OO
N
H
H
O
OH
OH
OH OH
O O
H3C
H3C CH3
H3CH3C
CH3
O
CH3
CH3
CH3
CH3
N(CH3)2
O
O
S
N

25.1 CLASSIFICATION OF NATURAL PRODUCTS 1017
WHY THIS CHAPTER?This small chapter merely tickles the surface of natural products chemistry,for hundreds, if not thousands, of books have been written on the subject.Rather than pretending to be comprehensive, this chapter is meant only toprovide a brief introduction to a large and immensely important area of modern biochemistry, perhaps tempting you to learn more on your own. Toprovide that introduction, we’ll look at the pathways by which several well-known natural products are synthesized in living organisms: pyridoxalphosphate (PLP), morphine, and erythromycin A. The molecules may appearcomplex (erythromycin A, in particular), but the individual chemical stepsby which they are made should be familiar to you at this point.
Classification of Natural ProductsThere is no rigid scheme for classifying natural products—their immensediversity in structure, function, and biosynthesis is too great to allow them to fit neatly into a few simple categories. In practice, however, workers in the field often speak of five main classes of natural products: terpenoids and steroids, fatty acid–derived substances and polyketides, alkaloids, non-ribosomal polypeptides, and enzyme cofactors.
• Terpenoids and steroids, as discussed previously in Chapter 23, are a vastgroup of substances—more than 35,000 are known—derived biosyntheti-cally from isopentenyl diphosphate. Terpenoids have an immense vari-ety of apparently unrelated structures, while steroids have a commontetracyclic carbon skeleton and are modified terpenoids that are biosyn-thesized from the triterpene lanosterol. We looked at terpenoid andsteroid biosynthesis in Sections 23.8–23.10.
• Alkaloids, like terpenoids, are a large and diverse class of compounds,with more than 12,000 examples known at present. They contain a basicamine group in their structure and are derived biosynthetically from aminoacids. We’ll look at morphine biosynthesis as an example in Section 25.3.
• Fatty acid–derived substances and polyketides, of which more than10,000 are known, are biosynthesized from simple acyl precursors suchas acetyl CoA, propionyl CoA, and methylmalonyl CoA. Natural products
Terpenoids,Steroids Alkaloids
Natural Products(secondary metabolites)
Fatty acids,Polyketides
Nonribosomalpolypeptides
Enzymecofactors
25.1

O HC
CH2OPO32–
OHH
OHH
D-Erythrose
4-phosphate
Pyruvate
D-Glyceraldehyde
3-phosphate
1 2 3
C O
C
C O
CH3
O O–C
CH2OPO32–
OHH
OHH
O O–C
CH2OPO32–
OPO32–
OHH
HH3N+
H3N+
D-Erythronate
4-phosphate
CH2OPO32–
OHH
HHO
C O
CH3
+CO2
–
1-Deoxyxylulose
5-phosphate
Pyridoxine
5�-phosphate
Pyridoxal
5�-phosphate (PLP)
O O–
CH2OPO32–
OHH
CO H
CH2OPO32–
OHH
3-Hydroxy-4-phospho-
hydroxy-2-ketobutyrate
4-Phospho-
hydroxythreonine
1-Amino-3-hydroxy-
acetone 3-phosphate
CO2CO2
4
5
6
O
CH2OPO32–
CH3
CH2OH
OHH+N
CH2OPO32–
CH3
CHO
OHH+N
7
1018 C H A P T E R 25: S E C O N DA RY M E TA B O L I T E S : A N I N T R O D U C T I O N TO N AT U R A L P R O D U C T S C H E M I S T RY
derived from fatty acids, such as the eicosanoid prostaglandin E1, gener-ally have most of the oxygen atoms removed, but polyketides, such as theantibiotic erythromycin A, often have many oxygen substituents remain-ing. We looked at eicosanoid biosynthesis as an example of a fattyacid–derived natural product in Section 23.7 and will look at erythro-mycin biosynthesis in Section 25.4.
• Nonribosomal polypeptides are peptidelike compounds that are biosyn-thesized from amino acids by a multifunctional enzyme complex withoutdirect RNA transcription. The penicillins are good examples, but theirchemistry is a bit complicated and we’ll not discuss their biosynthesis.
• Enzyme cofactors don’t fit one of the other general categories of naturalproducts and are usually classed separately. We’ve seen numerous exam-ples of coenzymes in past chapters (see the list in Table 19.3) and willlook at the biosynthesis of pyridoxal phosphate (PLP) in Section 25.2.
Figure 25.1 An overviewof the pathway for pyridoxal5�-phosphate biosynthesis.Individual steps areexplained in the text.

25.2 BIOSYNTHESIS OF PYRIDOXAL PHOSPHATE 1019
As you might imagine, unraveling the biosynthetic pathways by whichspecific natural products are made is extremely difficult and time-consumingwork. Small precursor molecules have to be identified, guesses about likelyroutes made, and individual enzymes that catalyze each step isolated, charac-terized, and mechanistically studied. The payoff for all this painstaking workis a fundamental understanding of how organisms function at the molecularlevel, an understanding that can be used to design new pharmaceutical agents.
Biosynthesis of Pyridoxal PhosphateLet’s begin this quick tour of natural products chemistry by looking at thebiosynthesis of pyridoxal 5�-phosphate (PLP), a relatively simple enzymecofactor we’ve encountered several times in different metabolic pathways.An overview of PLP biosynthesis is shown in Figure 25.1.
STEPS 1–2 OF FIGURE 25.1: OXIDATION Pyridoxal phosphate biosynthesis beginswith oxidation of the aldehyde group in D-erythrose 4-phosphate to give thecorresponding carboxylic acid, D-erythronate 4-phosphate. The oxidationrequires NAD� as cofactor and occurs by a mechanism similar to that of step 6in glycolysis, in which glyceraldehyde 3-phosphate is oxidized to the corre-sponding acid (see Section 22.2, Figure 22.6). A cysteine –SH group in theenzyme adds to the aldehyde carbonyl group of D-erythrose 4-phosphate to givean intermediate hemithioacetal, which is then oxidized by NAD� to a thioester.Hydrolysis of the thioester yields erythronate 4-phosphate, and a further oxi-dation of the –OH group at C2 by NAD� gives 3-hydroxy-4-phosphohydroxy-2-ketobutyrate (Figure 25.2).
O HC
CH2OPO32–
OHH
OHH
D-Erythrose
4-phosphate
CH2OPO32–
NAD+
OHH
OHH
Hemithioacetal
H
HA
O SC
CH2OPO32–
OHH
OHH
Thioester
EnzS
B
B
O O–C
CH2OPO32–
OHH
OHH
D-Erythronate
4-phosphate
CO HH
S
Enz
Enz
CONH2
N+
NADH/H+
NADH/H+
NAD+
C O
CO O–
CH2OPO32–
OHH
3-Hydroxy-4-phospho-
hydroxy-2-ketobutyrate
H2O Enz SH
25.2
Figure 25.2 Mechanism of steps 1and 2 in PLP biosynthesis, the oxida-tion of D-erythrose 4-phosphate togive 3-hydroxy-4-phosphohydroxy-2-ketobutyrate.

1020 C H A P T E R 25: S E C O N DA RY M E TA B O L I T E S : A N I N T R O D U C T I O N TO N AT U R A L P R O D U C T S C H E M I S T RY
STEPS 3–4 OF FIGURE 25.1: TRANSAMINATION AND OXIDATION/DECARBOXYLATION3-Hydroxy-4-phosphohydroxy-2-ketobutyrate undergoes a transamination instep 3 on reaction with �-ketoglutarate by the usual PLP-dependent mecha-nism, shown previously in Section 20.2, Figure 20.2. The product, 4-phospho-hydroxythreonine, is then oxidized by NAD� to give an intermediate �-ketoester, which undergoes concurrent decarboxylation and yields 1-amino-3-hydroxyacetone 3-phosphate. The reactions are shown in Figure 25.3.
STEP 5 OF FIGURE 25.1: FORMATION OF 1-DEOXYXYLULOSE 5-PHOSPHATE The 1-amino-3-hydroxyacetone 3-phosphate formed in step 4 of PLP biosynthesis reacts instep 6 with 1-deoxyxylulose 5-phosphate (DXP). DXP arises in step 5 by analdol-like condensation of D-glyceraldehyde 3-phosphate with pyruvate in athiamin-dependent reaction catalyzed by DXP synthase.
You might recall from Section 22.3, Figure 22.7, that pyruvate is con-verted to acetyl CoA by a process that begins with addition of thiamin diphos-phate (TPP) ylide to the ketone carbonyl group, followed by decarboxylationto give hydroxyethylthiamin diphosphate (HETPP). Exactly the same reac-tion occurs in DXP biosynthesis, but instead of reacting with lipoamide togive a thioester, as in the formation of acetyl CoA, HETPP adds to glycer-aldehyde 3-phosphate in an aldol-like reaction. The tetrahedral intermediatethat results expels TPP ylide as leaving group and yields DXP. The mecha-nism is shown in Figure 25.4.
STEP 6 OF FIGURE 25.1: CONDENSATION AND CYCLIZATION 1-Deoxy-D-xylulose 5-phosphate is dephosphorylated and then condenses with 1-amino-3-hydroxyacetone 3-phosphate in step 6 to give pyridoxine 5�-phosphate.The reaction begins with formation of an enamine, followed by loss of waterto form an enol that also contains a ketone group six atoms away. The enoladds to the ketone in an intramolecular aldol reaction (see Section 17.7) to
O O–C
CH2OPO32–
OHH
H
4-Phosphohydroxy-
threonine
NADH/H+
NAD+
C O
CO O–
CH2OPO32–
OHH
3-Hydroxy-4-phospho-
hydroxy-2-ketobutyrate
C O
CO O–
CH2OPO32–
H
A �-keto ester
H3N+Glutamate
�-Ketoglutarate
CO2
OPO32–H3N
+H3N
+
1-Amino-3-hydroxy-
acetone 3-phosphate
O
H A
Figure 25.3 Mechanism of steps 3and 4 in PLP biosynthesis.

25.2 BIOSYNTHESIS OF PYRIDOXAL PHOSPHATE 1021
Thiamin diphosphate ylide adds to theketone carbonyl group of pyruvate toyield an alcohol addition product.
1
The addition product contains a C=Nbond two carbons away from thecarboxylate and is structurally similarto a �-keto acid. It therefore loses CO2,giving the enamine HETPP.
2
The enamine adds to glyceraldehyde3-phosphate in an aldol-like reaction.
3
Cleavage of the adduct in a retro-aldolreaction gives 1-deoxy-D-xylulose5-phosphate and regenerates TPP ylide.
4
TPP ylide Pyruvate
H3C
H3C
CH3
R�
–
S
+
R
N
H3C
H3C
R� S
+
+
R
N
C
C
–O
–O
O
O
O
OH
H A
1
2
3
CO2
R�
R
S
N
H3C
H3C
H3C
R�
R
S
N
+
H3C
R�
R+
S
N
OH
CH3 H
H
OH
OPO32–
OPO32–
H OH
HHO
O
O
H
H
A
HETPP
Glyeraldehyde
3-phosphate
OPO32–
H OH
HHO
B
4
–
TPP ylide 1-Deoxy-D-xylulose
5-phosphate
O
Figure 25.4 M E C H A N I S M : Mechanism of step 5 in pyridoxal phosphate biosynthesis, the thiamin-dependent aldol reaction of D-glyceraldehyde 3-phosphate with pyruvate to give 1-deoxyxylulose5-phosphate.
© J
ohn
McM
urry

1022 C H A P T E R 25: S E C O N DA RY M E TA B O L I T E S : A N I N T R O D U C T I O N TO N AT U R A L P R O D U C T S C H E M I S T RY
form a six-membered ring, which then loses water. Tautomerization of theresultant unsaturated ketone gives an aromatic pyridine ring. Note that a lossof phosphate ion occurs at some point in the process, although the exact pointat which this happens is not known. The mechanism is shown in Figure 25.5.
Nucleophilic addition of the amineto 1-deoxy-D-xylulose gives anenamine . . .
1
. . . which loses water to form anenol that also contains a ketonegroup six atoms away.
2
The enol undergoes anintramolecular aldol reactionwith the ketone . . .
3
. . . and the aldol intermediate thenloses water. Tautomerization of thecarbonyl group yields pyridoxine5�-phosphate.
4
1-Deoxy-D-xylulose
5-phosphate
1-Amino-3-hydroxy-
acetone 3-phosphate
Enamine
Enol
CH2OPO32–
CH2OPO32–
CH2OPO32–
2–O3PO
CH3
CH3
H2N
O O
HO
HO
+
+ H2O
O
OH
OH
1
N
H
H
CH2OPO32–
CH2OPO32–
2–O3PO
2–O3PO
CH3
O
O HN
H
H
H
A
2
+
HA
HA
B
B
CH3
ON+
HO
3
4
CH2OPO32–
CH3
CH2OH
OHHN+
+ H2O + Pi
Pyridoxine
5�-phosphate
Figure 25.5M E C H A N I S M :Mechanism of step 6 inPLP biosynthesis, thereaction of 1-amino-3-hydroxyacetone 3-phosphate with 1-deoxy-D-xylulose 5-phosphate to givepyridoxine 5�-phosphate.
© J
ohn
McM
urry

25.3 BIOSYNTHESIS OF MORPHINE 1023
STEP 7 OF FIGURE 25.1: OXIDATION The final step in PLP biosynthesis is oxida-tion of the primary alcohol group in pyridoxine 5�-phosphate to the corre-sponding aldehyde. Typically, as we’ve seen on numerous occasions, alcoholoxidations are carried out by either NAD� or NADP�. In this instance, how-ever, flavin mononucleotide (FMN) is involved as the oxidizing coenzymeand reduced flavin mononucleotide (FMNH2) is the by-product. The detailsof the reaction are not clear, but evidence suggests that a hydride transfer isinvolved, just as in NAD� oxidations.
Problem 25.1 In the addition of HETPP to glyceraldehyde 3-phosphate shown in Figure25.4, does the reaction take place on the Re face or the Si face of the glycer-aldehyde carbonyl group?
Problem 25.2 Show a likely mechanism for the final tautomerization in the reaction of 1-amino-3-hydroxyacetone 3-phosphate with 1-deoxy-D-xylulose to givepyridoxine 5�-phosphate (Figure 25.5).
Biosynthesis of MorphineHaving looked at the biosynthesis of pyridoxal 5�-phosphate in the previoussection, let’s now go up a level in complexity by looking at morphine biosyn-thesis. Morphine, perhaps the oldest and best known of all alkaloids, isobtained from the opium poppy Papaver somniferum, which has been culti-vated for more than 6000 years. Medical uses of the poppy have been knownsince the early 1500s, when crude extracts, called opium, were used for therelief of pain. Morphine was the first pure compound to be isolated fromopium, but its close relative codeine also occurs naturally. Codeine, which issimply the methyl ether of morphine and is converted to morphine in the
25.3
Pyridoxine
5�-phosphate
Flavin mono-
nucleotide (FMN)
2–O3POCH2
CH3
O
OHH
H
+N
H
H
H B
H3C
H3C
O
O
N
N N
N
H A
Pyridoxal
5�-phosphate (PLP)
Reduced flavin mono-
nucleotide (FMNH2)
2–O3POCH2
CH3
O
OHH+N
H
H3C
H3C
O
O
N
H
N N
H
N
C
H

1024 C H A P T E R 25: S E C O N DA RY M E TA B O L I T E S : A N I N T R O D U C T I O N TO N AT U R A L P R O D U C T S C H E M I S T RY
body, is used in prescription cough medicines and as an analgesic. Heroin,another close relative of morphine, does not occur naturally but is synthe-sized in the laboratory by diacetylation of morphine.
Chemical investigations into the structure of morphine occupied some ofthe finest chemical minds of the 19th and early 20th centuries, and it was notuntil 1924 that the puzzle was finally solved by Robert Robinson, who receivedthe 1947 Nobel Prize in Chemistry for this and other work with alkaloids.
Morphine and its relatives are extremely useful pharmaceutical agents,yet they also pose an enormous social problem because of their addictiveproperties. Much effort has therefore gone into understanding how morphineworks and into developing modified morphine analogs that retain the anal-gesic activity but don’t cause physical dependence. Our present understand-ing is that morphine functions by binding to so-called mu opioid receptor sitesin both the spinal cord, where it interferes with the transmission of pain sig-nals, and brain neurons, where it changes the brain’s reception of the signal.
Hundreds of morphine-like molecules have been synthesized and testedfor their analgesic properties. Research has shown that not all the complexframework of morphine is necessary for biological activity. According to the “morphine rule,” biological activity requires (1) an aromatic ring attachedto (2) a quaternary carbon atom, followed by (3) two more carbon atoms and (4) a tertiary amine. Meperidine (Demerol), a widely used analgesic, andmethadone, a substance used in the treatment of heroin addiction, are twocompounds that fit the morphine rule.
The morphine rule
H
H
H H
HO
HO
O
CH3N
Methadone
CH3
H3C
An aromatic ringattached to a quaternary carbon ( )followed by two or more carbons ( )and a tertiary amine (N)
OCH2CH3
CH3
C6H5 N CH3NO
Meperidine
O
Morphine
H
H
H H H H
HO
HO
O
CH3N
Codeine
H
H
H
CH3O
HO
O
CH3N
Heroin
H
H
H
CH3CO
CH3CO
O
CH3N
O
O

25.3 BIOSYNTHESIS OF MORPHINE 1025
Morphine is biosynthesized from two molecules of the amino acid tyro-sine. One tyrosine is converted into dopamine, the second is converted intop-hydroxyphenylacetaldehyde, and the two are coupled to give morphine.The entire pathway is a bit complex at several points, but an abbreviatedscheme is given in Figure 25.6.
Figure 25.6 An abbreviated pathway for the biosynthesis of morphine from two molecules of tyrosine.The individual steps are explained in more detail in the text.
STEP 1 OF FIGURE 25.6: DOPAMINE BIOSYNTHESIS Dopamine is formed from tyro-sine in two steps: an initial hydroxylation of the aromatic ring, followed bydecarboxylation. The hydroxylation is catalyzed by tyrosine 3-mono-oxygenase, requires a cofactor called tetrahydrobiopterin, and occurs througha somewhat complex pathway that involves an iron–oxo (Fe�O) complexanalogous to that involved in prostaglandin biosynthesis (see Figure 7.9). The
Morphine
H
H
H H
HO
HO
O
CH3N
Codeine
H
H
H H
CH3O
HO
O
CH3N
Thebaine
HH
CH3O
CH3O
O
CH3N
Salutaridine(R)-Reticuline
H
CH3O
CH3O
HO
CH3N
O
H
CH3O
CH3O
HO
CH3N
OH
CH3O
CH3O
HO
=HO
CH3H
N
HO
HO
HOH
NH
6
(S)-Norcoclaurine
Dopamine
Tyrosine
4
3
2
1
7 8
5
HO
HO
HO
HO
CHO
NH2
p-Hydroxyphenyl-
acetaldehyde
CO2–
H3N+ H

1026 C H A P T E R 25: S E C O N DA RY M E TA B O L I T E S : A N I N T R O D U C T I O N TO N AT U R A L P R O D U C T S C H E M I S T RY
decarboxylation is catalyzed by the PLP-dependent enzyme aromatic L-amino acid decarboxylase.
Recall from Section 20.2 that pyridoxal 5�-phosphate reacts with the � amino group of an �-amino acid to form an imine, or Schiff base. When L-dopareacts with PLP, the resultant imine undergoes decarboxylation, with thepyridinium ion of PLP acting as the electron acceptor. Hydrolysis then givesdopamine and regenerated PLP. The mechanism is shown in Figure 25.7.
H
Pyridoxal
phosphate (PLP)
L-Dopa–PLP imineL-Dopa
2–O3PO
CH3
O
OH+N
C
H
H
HO
HO
+
2–O3PO
CH3
OH+N
N
H
H
H3N+ O–
C
O
H
HO
HO
O–C
O
2–O3PO
CH3
OHN
N
H
H
OH
OH
OH
OH
H
2–O3PO
CH3
OH+N
N
H
H H
HO
HO
H
CO2
H A
H2O
PLP Dopamine
2–O3PO
CH3
O
OH+N
C
H
H+
H3N+
L-Dopa
HO
HO CO2–
H
Dopamine
HO
HO NH3
CO2H2OO2
+
NH3+
Tyrosine
HO
CO2–
H NH3+
Figure 25.7Mechanism of step 1in morphine biosynthe-sis, the PLP-dependentdecarboxylation of L-dopa to givedopamine.
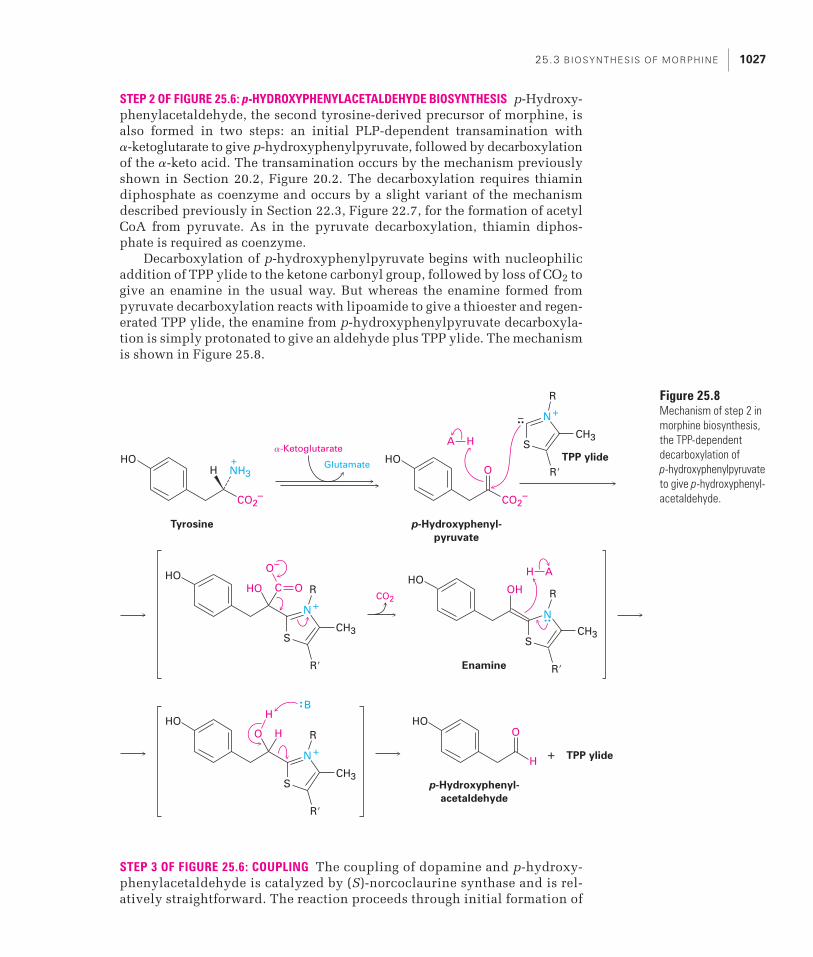
25.3 BIOSYNTHESIS OF MORPHINE 1027
STEP 2 OF FIGURE 25.6: p-HYDROXYPHENYLACETALDEHYDE BIOSYNTHESIS p-Hydroxy-phenylacetaldehyde, the second tyrosine-derived precursor of morphine, isalso formed in two steps: an initial PLP-dependent transamination with �-ketoglutarate to give p-hydroxyphenylpyruvate, followed by decarboxylationof the �-keto acid. The transamination occurs by the mechanism previouslyshown in Section 20.2, Figure 20.2. The decarboxylation requires thiamindiphosphate as coenzyme and occurs by a slight variant of the mechanismdescribed previously in Section 22.3, Figure 22.7, for the formation of acetylCoA from pyruvate. As in the pyruvate decarboxylation, thiamin diphos-phate is required as coenzyme.
Decarboxylation of p-hydroxyphenylpyruvate begins with nucleophilicaddition of TPP ylide to the ketone carbonyl group, followed by loss of CO2 togive an enamine in the usual way. But whereas the enamine formed frompyruvate decarboxylation reacts with lipoamide to give a thioester and regen-erated TPP ylide, the enamine from p-hydroxyphenylpyruvate decarboxyla-tion is simply protonated to give an aldehyde plus TPP ylide. The mechanismis shown in Figure 25.8.
STEP 3 OF FIGURE 25.6: COUPLING The coupling of dopamine and p-hydroxy-phenylacetaldehyde is catalyzed by (S)-norcoclaurine synthase and is rel-atively straightforward. The reaction proceeds through initial formation of
CO2
Tyrosine
HO
CO2–
H NH3+
p-Hydroxyphenyl-
pyruvate
p-Hydroxyphenyl-
acetaldehyde
Enamine
HO
CO2–
O–
CH3
Glutamate
�-Ketoglutarate
O
O
HOHOHO OH
HATPP ylide
R�
–
S
+
R
N
CH3
R�
S
R
N
H
H
A
C O
CH3
R�
S
+
R
N
+
HOO
HO
TPP ylide+
H
H
CH3
R�
S
R
N
B
Figure 25.8Mechanism of step 2 inmorphine biosynthesis,the TPP-dependentdecarboxylation of p-hydroxyphenylpyruvateto give p-hydroxyphenyl-acetaldehyde.

1028 C H A P T E R 25: S E C O N DA RY M E TA B O L I T E S : A N I N T R O D U C T I O N TO N AT U R A L P R O D U C T S C H E M I S T RY
an intermediate iminium ion, followed by intramolecular electrophilicaromatic substitution at a position para to one of the hydroxyl groups (Figure 25.9).
STEP 4 OF FIGURE 25.6: METHYLATION, HYDROXYLATION, AND EPIMERIZATION(S)-Norcoclaurine next undergoes two methylations and a hydroxylation to give(S)-3�-hydroxy-N-methylcoclaurine, which is methylated a third time to pro-duce (S)-reticuline. Epimerization of (S)-reticuline then yields (R)-reticuline(Figure 25.10).
Both initial methylations use S-adenosylmethionine (SAM) as the methyldonor, as discussed in Section 10.9. S-Adenosylhomocysteine (SAH) is theby-product in each case, and the reactions occur by the usual SN2 substitu-tion pathway. The first methylation occurs on a phenol oxygen, and the sec-ond takes place on the amine nitrogen.
The hydroxylation of (S)-N-methylcoclaurine to give (S)-3�-hydroxy-N-methylcoclaurine is superficially similar to the hydroxylation of tyrosine instep 1 in that both involve an iron–oxo complex as the active hydroxylatingagent. Unlike the enzyme in the tyrosine hydroxylation, however, thatresponsible for hydroxylation of N-methylcoclaurine is a so-called cyto-chrome P450 enzyme. These enzymes, of which more than 500 are known,contain an iron–heme cofactor ligated to the sulfur atom of a cysteine residuein the enzyme. The details of the hydroxylation itself are not clear, although
HO+
HO
HOH
NH
Dopamine
HO
HO
HOH
NH
(S)-Norcoclaurine
HO
HO
HO
CHO H
HO
NH2
HO
HO
p-Hydroxyphenyl-
acetaldehyde
Iminium ion
H2O NH+
B
Figure 25.9 Mechanismof step 3 in morphinebiosynthesis, the coupling of dopamine and p-hydroxyphenyl-acetaldehyde to give (S)-norcoclaurine.

25.3 BIOSYNTHESIS OF MORPHINE 1029
it may occur through a straightforward electrophilic aromatic substitutionmechanism.
Methylation of a phenolic –OH group in (S)-3�-hydroxy-N-methyl-coclaurine by SAM gives (S)-reticuline through the usual SN2 pathway, and epimerization of the chirality center forms (R)-reticuline. The epimer-ization is a two-step process, the first an oxidation of the tertiary amine to an intermediate iminium ion and the second a hydride reduction of
CH3
CH3
H3C
H3C
Heme
HO2C CO2H
Fe(II)
N N
N N
CH3
CH3
H3C
H3C
Heme iron–oxo complex
HO2C CO2H
Fe(V)
N N
N N
O2
O
S
Cys
Enz
HO
HO
HOH
NH
(S)-Norcoclaurine
CH3O
HO
HOH
NH
(S)-Coclaurine
SAHSAM SAHSAM O2 H2O
CH3O
HO
HOH
N
(S)-N-Methylcoclaurine
CH3
SAHSAM
CH3O
HO
HO
HOH
N
(S)-3�-Hydroxy-N-
methylcoclaurine
CH3
CH3O
CH3O
HO
HOH
N
(S)-Reticuline
CH3
CH3O
CH3O
HO
HON
(R)-Reticuline
CH3H
Figure 25.10 An overview of the reactions in step 4 of morphine biosynthesis, the conversion of (S)-norcoclaurine to (R)-reticuline.

1030 C H A P T E R 25: S E C O N DA RY M E TA B O L I T E S : A N I N T R O D U C T I O N TO N AT U R A L P R O D U C T S C H E M I S T RY
the iminium ion. The mechanism of the oxidation step is not yet known,but the reduction of the iminium ion requires NADPH as cofactor (Fig-ure 25.11).
Why does morphine biosynthesis proceed through initial formation of(S)-reticuline as an intermediate, followed by epimerization, rather thanthrough (R)-reticuline directly? There is no obvious answer other than tosay that many metabolic pathways contain such small inefficiencies, prob-ably as a result of the evolutionary development of the responsibleenzymes—what some people have called “unintelligent design.”
STEP 5 OF FIGURE 25.6: OXIDATIVE COUPLING (R)-Reticuline is converted intosalutaridine in step 5 by an oxidative coupling between the ortho positionof one phenol ring and the para position of the other. The reaction is cat-alyzed by another cytochrome P450 enzyme like that involved in thehydroxylation of (S)-N-methylcoclaurine in step 4. Formation of the phen-oxide ions and abstraction of a nonbonding electron from each oxygen atomto give radicals occurs, followed by radical coupling and a keto–enol tau-tomerization to yield salutaridine (Figure 25.12).
STEP 6 OF FIGURE 25.6: REDUCTION AND CYCLIZATION Reduction of salutaridineto salutaridinol is catalyzed by salutaridine reductase, with NADPH ascofactor. This alcohol then undergoes a nucleophilic acyl substitutionreaction with acetyl CoA to give a doubly allylic acetate, which sponta-neously eliminates acetate ion in an SN1-like process and cyclizes to the-baine (Figure 25.13).
CH3O
CH3O
HO
HOH
N
(S)-Reticuline
CH3
CH3O
CH3O
HO
HON+
Iminium ion
CH3
(R)-Reticuline
H
CH3O
CH3O
HO
CH3N
OH
CH3O
CH3O
HO
=HO
CH3H
NNADP+
NADPH/H+
Figure 25.11 Mechanism of theepimerization of (S)-reticuline to (R)-reticuline in step 4 of morphinebiosynthesis.

25.3 BIOSYNTHESIS OF MORPHINE 1031
Salutaridine
H
CH3O
O
CH3O
HO
Salutaridinol
H
CH3O
CH3CSCoA
HSCoA
CH3O
HONADP+
NADPH/H+
H OH
H
CH3O
CH3O
HO
H OCOCH3
O
H
H
+CH3O
CH3O
OHCH3CO2–
B
Thebaine
HH
CH3O
CH3O
O
NCH3
NCH3
NCH3
NCH3
NCH3
(R)-Reticuline
Salutaridine
H
CH3O
CH3O
HO
CH3N
OH
H
H
CH3O
CH3O
•O
CH3N
O•
•
•
H
CH3O
CH3O
O
O
CH3N
H
H
CH3O
CH3O
O
O
CH3NH
CH3O
O
CH3N
CH3O
HO
Figure 25.12 Mechanism of step 5 in morphine biosynthesis, the oxidative phenol coupling of (R)-reticuline to salutaridine.
Figure 25.13 Mechanism ofstep 6 in morphine biosynthesis,the formation of thebaine fromsalutaridine.

1032 C H A P T E R 25: S E C O N DA RY M E TA B O L I T E S : A N I N T R O D U C T I O N TO N AT U R A L P R O D U C T S C H E M I S T RY
STEPS 7–8 OF FIGURE 25.6: DEMETHYLATION AND REDUCTION The remaining stepsin the biosynthesis of morphine involve two demethylation reactions and areduction. The first demethylation is catalyzed by a cytochrome P450enzyme, which hydroxylates the –OCH3 group of thebaine to form –OCH2OH,a hemiacetal. Loss of formaldehyde then gives an enol that tautomerizes tocodeinone. Reduction of the resultant ketone by NADPH yields codeine, anddemethylation by a P450 enzyme produces morphine (Figure 25.14).
Problem 25.3 Show the mechanism of the reaction of (S)-norcoclaurine with S-adenosyl-methionine to give (S)-coclaurine (Figure 25.10).
Morphine
H
H
H
HO
HO
O
CH3N
Codeine
H
H
H
CH3O
HO
O
CH3N
Codeinone
HH
CH3O
O
CH3N
Thebaine
HH
CH3O
CH3
O
O
CH3N HH
CH3O
CH2•
O
O
CH3N
NADP+
NADPH/H+
HH
CH3O
CH2
O
O
CH3N
H
OH A
B
CH2O
O
O2H2OCH2O,
O2H2O
H
HH
Figure 25.14 Mechanism of step 7in morphine biosynthesis, the demethyl-ation of thebaine to give codeinone,catalyzed by a P450 enzyme. Reductionof codeinone with NADPH then yieldscodeine, and a final demethylation pro-duces morphine.

25.4 BIOSYNTHESIS OF ERYTHROMYCIN 1033
Problem 25.4 Convince yourself that the following two structures both represent (R)-reticuline.Which carbon atoms in the structure on the right correspond to the two carbonsindicated in the structure on the left?
Biosynthesis of ErythromycinHaving discussed the biosynthesis of pyridoxal phosphate and morphine in thepreceding two sections, we’ll end this chapter on natural products chemistryby going up yet one more level in complexity and looking at polyketide biosyn-thesis. Unlike what happens in many metabolic pathways, where each separatestep is catalyzed by a separate, relatively small enzyme, erythromycin andother polyketides are assembled by a single massive enzyme called a synthase.The synthase contains many enzyme domains linked together, with eachdomain catalyzing a specific biosynthetic step in sequence.
Polyketides are an extraordinarily valuable class of natural productsnumbering over 10,000 compounds. Commercially important polyketidesinclude antibiotics (erythromycin A, tetracycline) and immunosuppressants(rapamycin), as well as anticancer (doxorubicin), antifungal (amphotericinB), and cholesterol-lowering (lovastatin) agents (Figure 25.15). It has beenestimated that the sales of these and other polyketide pharmaceuticals totalmore than $15 billion per year.
Polyketides are biosynthesized by the joining together of the simple acylCoA’s acetyl CoA, propionyl CoA, methylmalonyl CoA, and (less frequently)butyryl CoA. The key carbon–carbon bond-forming step in each joining is aClaisen condensation (see Section 17.8). Once the carbon chain is assembledand released from the enzyme, further transformations take place to give thefinal product. Erythromycin A, for instance, is prepared from one propionateand six methylmalonate units by the pathway outlined in Figure 25.16. Following initial assembly of the acyl units into the macrocyclic lactone 6-deoxyerythronolide B, two hydroxylations, two glycosylations, and a finalmethylation complete the biosynthesis.
The initial assembly of seven acyl CoA precursors to build a poly-ketide carbon chain is carried out by a multienzyme complex called a polyketide synthase, or PKS. The 6-deoxyerythronolide B synthase (DEBS)is a massive structure of greater than 2 million molecular weight that contains more than 20,000 amino acids. Furthermore, it is a homodimer,
25.4
H
CH3O
CH3O
HO
CH3N
OH
CH3O
CH3O
HO
=HO
CH3H
N

1034 C H A P T E R 25: S E C O N DA RY M E TA B O L I T E S : A N I N T R O D U C T I O N TO N AT U R A L P R O D U C T S C H E M I S T RY
H
H
H
OH
OH
O
O O
O
O
CH3
NHH
OHOH
OH
Tetracycline
(antibiotic)
OH
O O O
NH2
H3C OHCH3H3C
OCH3
CH3 CH3
CH3
HO
OH
HO
O
O
OH
OH
Doxorubicin
(anticancer)
Rapamycin
(immunosuppressant)
Lovastatin
(cholesterol lowering)
Amphotericin B
(antifungal)
O
O O
CH2OH
H3C
NH2
O
OCH3CH3
H3C
H3C
H3C
CH3O
OH
CH3O
N
O
O
OH
CH3
CH3
H
H
OHOH
OH
H3C
H3C
H3C
O
O
O OHHO OH
O
OH OH
O
HONH2
OHO CH3
OCO2H
H
Figure 25.15 Structures of some polyketides used as pharmaceutical agents.

25.4 BIOSYNTHESIS OF ERYTHROMYCIN 1035
O
CH3
CH3
Erythronolide B6-Deoxyerythronolide BMethylmalonyl CoA
Propionyl CoA
CH3
H3C
H3C
OH
OH
H3C
OH
H3C
OH
O
O
O
CH3
CH3
CH3
H3C
H3C
OHH3C
OH
H3C
OH
O
OO
CoAS
+
CO2–
6
O
CoAS
O
CH3
CH3
3-O-Mycarosyl-
erythronolide B
CH3
H3C
H3C
OH
OH
H3C
OH
H3C
O
O
O
OH
OH
O
CH3
CH3
O
CH3
CH3
Erythromycin D
CH3
H3C
H3COH
H3C
OH
H3C
O
O
O
OH
OH
O
CH3
CH3
HOO
N(CH3)2
O CH3
O
CH3
CH3
Erythromycin C
CH3
H3C
H3COH
H3C
OH
OH
H3C
O
O
O
OH
OH
O
CH3
CH3
HOO
N(CH3)2
O CH3
O
CH3
CH3
Erythromycin A
CH3
H3C
H3COH
H3C
OH
OH
H3C
O
O
O
OH
OCH3
O
CH3
CH3
HOO
N(CH3)2
O CH3
Figure 25.16 An outline of the pathway for the biosynthesis of erythromycin A. One propionate and sixmethylmalonate units are first assembled into the macrocyclic lactone 6-deoxyerythronolide B, which isthen hydroxylated, glycosylated by two different sugars, hydroxylated again, and finally methylated.

1036 C H A P T E R 25: S E C O N DA RY M E TA B O L I T E S : A N I N T R O D U C T I O N TO N AT U R A L P R O D U C T S C H E M I S T RY
meaning that it consists of two identical protein chains held together bynoncovalent interactions, with each chain containing all the enzymes nec-essary for constructing the polyketide.
Each separate enzyme domain in the erythromycin synthase is a folded,globular region within a huge protein chain that catalyzes a specific biosyn-thetic step. The domains are grouped into modules, where each module car-ries out the sequential addition and processing of an acyl CoA to the growingpolyketide. In addition, adjacent modules form three larger groups (DEBS 1,DEBS 2, and DEBS 3) that are linked by peptide spacers. As shown in Figure25.17, the erythromycin PKS consists of an initial loading module to attachthe first acyl group, six extension modules to add six further acyl groups, andan ending module to cleave the thioester bond and release the polyketide.The ending module also catalyzes cyclization to give a macrocyclic lactone.
The loading module has two domains: an acyl transfer (AT) domain andan acyl carrier protein (ACP) domain. The AT selects the first acyl CoA (pro-pionyl CoA in the case of erythromycin) and transfers it to the adjacent ACP,which binds it through a thioester linkage and holds it for further reaction.Each extension module has a minimum of three domains: an AT, an ACP, anda ketosynthase (KS), which catalyzes the Claisen condensation reaction thatbuilds the polyketide chain. In addition to the three minimum domains, someextension modules also contain a ketoreductase (KR) to reduce a ketonecarbonyl group and produce an alcohol, a dehydratase (DH) to dehydrate thealcohol and produce a C�C bond, and an enoyl reductase (ER) to reduce the C�C bond. Finally, the ending domain is a thioesterase (TE), whichreleases the product by catalyzing a lactonization.
Polyketide chain extension occurs when an extension module AT selectsa new acyl CoA, transfers it to the ACP, and the KS then catalyzes a Claisencondensation reaction between the newly bonded acyl group and the acylgroup of the previous module. Figure 25.18 shows the steps occurring in thefirst extension cycle; other extension cycles take place similarly.
STEP 1 OF FIGURE 25.18: LOADING The loading AT domain begins the erythro-mycin biosynthesis by binding a propionyl CoA through a thioester bond tothe –SH of a cysteine residue. The AT then transfers the propionyl group tothe adjacent ACP. Each ACP in the synthase contains a phosphopantetheinebonded to the hydroxyl of a serine residue, and bonding of the acyl group tothe enzyme occurs by thioester formation with the phosphopantetheine –SH(Figure 25.19). The phosphopantetheine effectively acts as a long, flexiblearm to allow movement of the acyl group from one catalytic domain toanother.
STEPS 2–4 OF FIGURE 25.18: CHAIN EXTENSION Polyketide chain extension begins(step 2) when the acyl ACP of the loading module transfers the propionylgroup to the ketosynthase of module 1 (KS1), again forming a thioester bondto a cysteine residue. At the same time (step 3), the AT and ACP of module 1 load a (2S)-methylmalonyl CoA onto the thiol terminus of the ACP1 phos-phopantetheine. The key carbon–carbon bond formation occurs (step 4)when KS1 catalyzes a Claisen condensation and decarboxylation to form an

25.4 BIOSYNTHESIS OF ERYTHROMYCIN 1037
AC
PK
SK
SA
TA
TT
E
En
d
KR
Ex
ten
sio
n
Mo
du
le 6
DE
BS
3 (
3179 a
a)
Ex
ten
sio
n
Mo
du
le 5
KR
O O OH
OH
S
OH
OH
Hep
taketi
de
Hexaketi
de
O O OH
OH
S
OH
AC
PK
SA
TK
SD
HA
TA
CP
ER
KR
Exte
nsio
n
Mo
du
le 4
DE
BS
2 (
3568 a
a)
Exte
nsio
n
Mo
du
le 3 A
CP
O O OH
S
OH
Pen
taketi
de
O O OH
S
OH
Tetr
aketi
de
KR
AC
PK
SK
SA
CP
--
AT
KR
Exte
nsio
n
Mo
du
le 2
DE
BS
1 (
~3174 a
a)
Exte
nsio
n
Mo
du
le 1
Lo
ad
AT
AT
O
S
OH
Tri
keti
deO
H
AC
P
O
S
OH
Dik
eti
de
O
S
6-D
eo
xyery
thro
no
lid
e B
DE
BS
—6-
Deo
xyer
yth
ron
olid
e B
syn
thas
eA
T—
Acy
ltra
nsf
eras
eA
CP
—A
cyl c
arri
er p
rote
inK
S—
Ket
oac
yl s
ynth
ase
KR
—K
eto
acyl
red
uct
ase
DH
—D
ehyd
rata
seE
R—
En
oyl
red
uct
ase
TE
—T
hio
este
rase
O
CH
3 CH
3
CH
3
H3C
H3C
OH
H3C
OH
H3C
OH
O
O
Figu
re 2
5.17
A sc
hem
atic
vie
w o
f the
6-d
eoxy
eryt
hron
olid
e B
synt
hase
(DEB
S), s
how
ing
the
loca
tions
of t
he e
nzym
e do
mai
ns w
ithin
the
load
ing
mod
ule
and
the
six e
xten
sion
mod
ules
. The
figu
re is
exp
lain
ed in
det
ail in
the
text
.

1038 C H A P T E R 25: S E C O N DA RY M E TA B O L I T E S : A N I N T R O D U C T I O N TO N AT U R A L P R O D U C T S C H E M I S T RY
enzyme-bound �-keto thioester. It’s likely that the decarboxylation occurssimultaneously with the Claisen condensation, giving the enolate ion neces-sary for nucleophilic addition to the second thioester.
O
HS
AT
S
KS KR ACP
S
O
S
O–O
HS
AT
HS
KS KR ACP
S
O
R
O
CO2
Abbreviatedmechanism
Loadingmodule
Extensionmodule 1
O
S
AT
HS
AT
HS
KS
HS
ACP KR
HS
ACP
Loadingmodule
Extensionmodule 1
HS
O
S
AT
HS
AT
HS
KSACP KR
HS
ACP
HS
O
AT
HS
AT
S
KS
HS
ACP KR
HS
ACP
HS
O
AT
HS
AT
S
KS
HS
ACP KR ACP
HS
AT
HS
AT
HS
KS
HS
ACP KR
S
ACP
HS
AT
HS
AT
HS
KS
HS
ACP KR ACP
O
R
O
S
H3C
HS
AT
HS
AT
HS
KS
HS
ACP KR
S
ACP
O
S
O
H3C
S
O
S
O–O
S
O
OH
1 2
3 4
65
Figure 25.18 The initialloading and first chain-extension cycle catalyzedby the erythromycin PKS.Individual steps areexplained in the text.

25.4 BIOSYNTHESIS OF ERYTHROMYCIN 1039
STEPS 5–6 OF FIGURE 25.18: EPIMERIZATION AND REDUCTION Interestingly, the Claisencondensation occurs with inversion of configuration at the methyl-bearing chirality center so that the initially formed diketide has (R) stereochemistry.Base catalyzed epimerization of the (R) product, an acidic �-diketone, occurs instep 5, however, so the product that goes on to the next step regains the (S) con-figuration. Finally, KR1 reduces the ketone to a �-hydroxy thioester in step 6 by transfer of the pro-S hydrogen from NADPH as cofactor. Module 1 is now finished, so the diketide is transferred to KS2 for another chain extension.
The reactions catalyzed by extension modules 2, 5, and 6 are similar tothose of module 1, although the stereochemistries of the Claisen condensa-tion and reduction steps may differ. The reactions in modules 3 and 4, how-ever, are different. Module 3 lacks a KR domain, so no reduction occurs andthe tetraketide product contains a ketone carbonyl group (Figure 25.17). Mod-ule 4 contains a KR and two additional enzyme domains, so it catalyzes aketone reduction plus two additional reactions. Following the reduction byKR4 of the pentaketide, a dehydratase (DH) dehydrates the pentaketide alco-hol to an �,�-unsaturated thioester and the double bond is then reduced by anenoyl reductase (ER) domain (Figure 25.20).
HS
AT
HS
KS KR
S
ACP
HS
AT
HS
KS KR ACP
O
R
O
SH3C
HS
AT
HS
KS KR
S
ACP
O
S
O
H3C
S
O
OH
Abbreviatedmechanism
AT
ACP
O
S
AT
SH
B
Phosphopantetheine
Acyl ACP
O
CH2CH2NHCCH2CH2NHCCHCCH2OP
O
OH S
CH3 O–
OCH3
HO
ACP
O
CH2CH2NHCCH2CH2NHCCHCCH2OP
O
O=
S
CH3 O–
OCH3
HO
ACP
O
S
O
Figure 25.19 Formation of an acylACP during polyketide biosynthesis.Phosphopantetheine, symbolized by azigzag line between S and ACP, actsas a long, flexible arm to allow theacyl group to move from one catalyticdomain to another.

1040 C H A P T E R 25: S E C O N DA RY M E TA B O L I T E S : A N I N T R O D U C T I O N TO N AT U R A L P R O D U C T S C H E M I S T RY
Note that the complete sequence of reactions carried out by module 4—Claisen condensation, ketone reduction, dehydration, and double-bond reduction—is identical to the series of reactions found in fatty-acid biosynthesis(see Section 23.6, Figure 23.6). In fact, all fatty-acid synthases have the same setof AT, ACP, KS, KR, DH, and ER domains as the polyketide synthases.
Release of 6-deoxyerythronolide B from the PKS is catalyzed by the end-ing thioesterase module. A serine residue on the TE module first carries out anucleophilic acyl substitution on the ACP-bound heptaketide, and the acylenzyme that results undergoes lactonization. A histidine residue in the TEacts as base to catalyze nucleophilic acyl substitution of the serine ester bythe terminal –OH group in the heptaketide (Figure 25.21).
O
O
OH
OH
OH
OH
Heptaketide
6-Deoxyerythronolide B
O
CH3
CH3
CH3
H3C
H3C
OHH3C
OH
H3C
OH
O
O
S
O
O
O H
OH
OH
OH
O
OH
SerACP H A
TE
Ser TE
His
OHOH
A pentaketide
O
O
OH
OH
S
ACP
KR
OH
O
O
O
OH
S
ACP
O
O
OH
S
ACP
OH
O
O
OH
S
ACP
DH ER
Figure 25.20 Additional processingof the pentaketide intermediate inmodule 4 removes a carbonyl groupby a reduction–dehydration–reductionsequence.
Figure 25.21 Release of 6-deoxyerythronolide from the PKS occurs by lactonization of an acyl enzyme, formed by reaction of a serine residue in the TE modulewith the heptaketide.

25.4 BIOSYNTHESIS OF ERYTHROMYCIN 1041
Following its release from the PKS, 6-deoxyerythronolide B is hydroxyl-ated at C6 with retention of configuration to give erythronolide B. The reactionis catalyzed by a P450 hydroxylase analogous to that involved in morphinebiosynthesis (Section 25.3, Figure 25.14). L-Mycarose is then attached to the C3 hydroxyl group by reaction with thymidyl diphosphomycarose through anSN1-like process that proceeds by initial formation of the mycarosyl carbo-cation (Figure 25.22).
The final steps in erythromycin A biosynthesis are a further glycosyla-tion, a further hydroxylation, and a methylation (Figure 25.23). As in theattachment of mycarose, the attachment of the amino sugar D-desosaminealso takes place by transfer from a thymidyl diphosphosugar. C12 hydroxyla-tion by another P450 enzyme occurs with retention of configuration to giveerythromycin C, and methylation of the C3�� hydroxyl group of the mycaroseunit by reaction with S-adenosylmethionine gives erythromycin A.
Thymidine
6-Deoxyerythronolide B
O
CH3
CH3
CH3
H3C
H3C
OHH3C
OH
H3C
OH
O
O
O
CH3
CH3
Erythronolide B
CH3
H3C
H3C
OH
OH
H3C
OH
H3C
OH
O
O
O
CH3
CH3
3-O-Mycarosyl-
erythronolide B
CH3
H3C
H3C
OH
OH
H3C
OH
H3C
O
O
O
OH
OH
O
CH3
CH3
O26
3
O P O P
O–
O
O–
O
O
TDP
OH
OH
O
CH3
CH3
Figure 25.22 Hydroxylation and glycosylation of 6-deoxyerythronolide B to give 3-O-mycarosylerythronolide B.

1042 C H A P T E R 25: S E C O N DA RY M E TA B O L I T E S : A N I N T R O D U C T I O N TO N AT U R A L P R O D U C T S C H E M I S T RY
Problem 25.5 Show a likely mechanism for the epimerization that occurs in step 5 of Figure 25.18.
Problem 25.6 Propose a mechanism for the reaction of erythronolide B with thymidyldiphosphomycarose to give 3-O-mycarosylerythronolide B (Figure 25.22).
S
O
R
O
H3C
S
O
S
O
H3C
ACP ACP
O
CH3
CH3
3-O-Mycarosyl-
erythronolide B
CH3
H3C
H3C
OH
OH
H3C
OH
H3C
O
O
O
OH
OH
O
CH3
CH3
OTDP
TDP
O
CH3
CH3
Erythromycin D
CH3
H3C
H3COH
H3C
OH
H3C
O
O
O
OH
OH
O
CH3
CH3
HOO
N(CH3)2
O CH3
O
CH3
CH3
Erythromycin C
CH3
H3C
H3COH
H3C
OH
OH
H3C
O
O
O
OH
OH
O
CH3
CH3
HOO
N(CH3)2
O CH3
O
CH3
CH3
Erythromycin A
CH3
H3C
H3COH
H3C
OH
OH
H3C
O
O
O
OH
OCH3
O
CH3
CH3
HOO
N(CH3)2
O CH3
HO
N(CH3)2
O CH3
SAHSAM
O2
Figure 25.23 Final steps in the biosynthesis of erythromycin A.

SUMMARY 1043
SummaryThe term natural product is generally taken to mean a secondary metabolite—a small molecule that is not essential to the growth and development of theproducing organism and is not classified by structure. Well over 300,000 sec-ondary metabolites probably exist, generally classified into five categories:terpenoids and steroids, fatty acid–derived substances and polyketides,alkaloids, nonribosomal polypeptides, and enzyme cofactors.
Unraveling the biosynthetic pathways by which natural products aremade is difficult and time-consuming work, but the payoff is a fundamentalunderstanding of how organisms function at the molecular level. The mole-cules are sometimes complex, but the individual chemical steps by whichthey are made are familiar.
alkaloid, 1017
enzyme cofactor, 1018
fatty acid–derived substance,
1017
natural product, 1016
nonribosomal polypeptide,
1018
polyketide, 1017
secondary metabolite, 1016
steroid, 1017
terpenoid, 1017
Most chemists and biologistsspend the majority of theirtime in the laboratory. A few,however, spend their daysscuba diving on South Pacificislands or trekking throughthe rain forests of South Amer-ica and Southeast Asia. Theyaren’t on vacation, though;they’re at work as bioprospec-tors, and their job is to hunt fornew and unusual naturalproducts that might be usefulas drugs.
As noted in the Chapter 5Lagniappe, more than half ofall new drug candidates comeeither directly or indirectlyfrom natural products. Allfour natural products shownin the introduction to thischapter, for instance, are usedas drugs: morphine from theopium poppy, prostaglandinE1 from sheep prostate glands,
erythromycin A from a Streptomyces erythreus bacteriumcultured from a Philippine soil sample, and benzylpeni-cillin from Penicillium notatum. Still other examplesinclude rapamycin (Figure 25.15), an immunosuppressantisolated from a Streptomyces hygroscopicus bacterium first
found in a soil sample from Easter Island (Rapa Nui), andpaclitaxel (Taxol), an anticancer drug isolated from the barkof the Pacific yew tree found in the American Northwest.
With less than 1% of living organisms yet investigated,bioprospectors have a lot of work to do, but there is a racegoing on. Rain forests throughout the world are beingdestroyed at an alarming rate, causing many species ofboth plants and animals to become extinct before they caneven be examined. Fortunately, the governments in manycountries seem aware of the problem, but there is as yet nointernational treaty on biodiversity that could help pre-serve vanishing species.
O
O
O
O
Paclitaxel (Taxol)
OO
O
OO
N
H
H
OH CH3
OH
OH
O
H3C
H
O
Lagniappe
Bioprospecting: Hunting for Natural Products
Rapamycin, an immunosup-pressant natural product usedduring organ transplants, wasoriginally isolated from a soilsample found on EasterIsland, or Rapa Nui, an island2200 miles off the coast ofChile known for its giant Moaistatues.
© R
oyal
ty-fr
ee/C
orbi
s

1044 C H A P T E R 25: S E C O N DA RY M E TA B O L I T E S : A N I N T R O D U C T I O N TO N AT U R A L P R O D U C T S C H E M I S T RY
Exercises
Assess your understanding of this chapter’s topics with additional quizzing and conceptually based problems at http://now.brookscole.com/mcmurryorgbio1
25.7 Which hydrogen, pro-R or pro-S, is removed from pyridoxine 5�-phos-phate in the final step of PLP biosynthesis?
25.8 Does the ketone reduction step catalyzed by KR1 in erythromycinbiosynthesis occur on the Re or the Si face of the substrate carbonylgroup? (See Figure 25.18.)
25.9 When the enoyl reductase domain (ER4) in the erythromycin PKS isdeactivated by gene mutation, all further steps still occur normally.What is the structure of the lactone that results?
25.10 One of the steps in the biosynthesis of the alkaloid berbamunine is anepimerization of (S)-N-methylcoclaurine. Review the morphine biosyn-thesis in Figure 25.6, and propose a mechanism for the epimerization.
25.11 The final step in the biosynthesis of berbamunine is a coupling reactionof (S)-N-methylcoclaurine with (R)-N-methylcoclaurine (Problem 25.10).Propose a mechanism.
CH3O
OH
O
HOH
NCH3
Berbamunine
CH3O
HON
CH3H
CH3O
HO
HOH
N
(S)-N-Methylcoclaurine
CH3
CH3O
HO
HON
(R)-N-Methylcoclaurine
CH3H
Pyridoxine
5�-phosphate
2–O3POCH2
CH3
OH
OHH+N
HH
Pyridoxal
5�-phosphate (PLP)
2–O3POCH2
CH3
O
OHH
H
+N
FMNH2FMN

EXERCISES 1045
25.12 5-Aminolevulinate is the precursor from which the large class of alkaloids called tetrapyrroles are biosynthesized. It arises by a PLP-dependent reaction of glycine and succinyl CoA. Review themechanism of the formation of dopamine from L-dopa in Figure 25.7,and propose a mechanism for 5-aminolevulinate biosynthesis.
25.13 One of the steps in the biosynthesis of penicillins is a PLP-dependentepimerization of isopenicillin N to penicillin N.
The reaction occurs by initial formation of an imine, followed by abase-catalyzed isomerization. Propose a mechanism.
25.14 Propose a mechanism for the following biosynthetic conversion. Whatcofactors are likely to be involved?
25.15 The enzyme acetolactate synthase catalyzes the thiamin-dependentconversion of two molecules of pyruvate to acetolactate. Propose amechanism.
H3C
CH3HO
CO2–
O
H3C2
CO2–
O CO2
HN+
O
ONH
CO2–
CO2CH3CO2
–
Penicillin N
H H
CH3
CH3
CO2–
–O2C
OO
N
H
H
S
NHH3N+
(PLP)
Isopenicillin N
H H
CH3
CH3
CO2–
–O2C
OO
N
H
H
S
NH NH3+
5-AminolevulinateSuccinyl CoAGlycine
H3N CO2–
CO2–CoAS
++H2N
+O
CO2–
O(PLP)

1046 C H A P T E R 25: S E C O N DA RY M E TA B O L I T E S : A N I N T R O D U C T I O N TO N AT U R A L P R O D U C T S C H E M I S T RY
25.16 1-Deoxy-D-xylulose 5-phosphate (DXP), in addition to being a precur-sor to PLP, is also a precursor to isopentenyl diphosphate in terpenoidbiosynthesis. The initial step in the pathway is a base-catalyzedrearrangement, followed by reduction with NADPH to give 2C-methyl-D-erythritol 4-phosphate. Show the structure of the rearranged inter-mediate, and propose a mechanism for its formation.
25.17 Biosynthesis of the �-lactam antibiotic clavulanic acid begins with aTPP-dependent reaction between D-glyceraldehyde 3-phosphate andarginine.
(a) The first step is the reaction of D-glyceraldehyde 3-phosphate withTPP ylide, followed by dehydration to give an enol. Show themechanism, and draw the structure of the product.
(b) The second step is loss of hydrogen phosphate from the enol to givean unsaturated carbonyl compound. Show the mechanism, anddraw the structure of the product.
(c) The third step is a conjugate of arginine to the unsaturatedcarbonyl compound. Show the mechanism, and draw the structureof the product.
(d) The final step is a base-catalyzed hydrolysis to give the final prod-uct and regenerate TPP ylide. Show the mechanism.
H CO2–
Arginine
Clavulanic acid
HO
OHC
H
D-Glyceraldehyde
3-phosphate
OPO32– H2N+
N
H
NH2+
NH2
H CO2–
–O2C N
H
(TPP)
N
H
H
CO2–
CH2OH
H
NO
NH2+
NH2
O
H3C
H
HO
HO
H
OPO32–
H3C OH
HO
HO
HH
1-Deoxy-D-xylulose
5-phosphate
2C-Methyl-D-erythritol
4-phosphate
H
OPO32–[ ? ]
NADPH/H+ NADP+