2 is a missing link in plant cell ca2 signalling and
TRANSCRIPT

International Journal of
Molecular Sciences
Article
Mg2+ is a Missing Link in Plant Cell Ca2+ Signallingand Homeostasis—A Study on Vicia faba Guard Cells
Fouad Lemtiri-Chlieh 1,2,*, Stefan T. Arold 1,3,4 and Chris Gehring 1,5,*1 King Abdullah University of Science and Technology (KAUST), Biological and Environmental Science and
Engineering (BESE), Thuwal 23955-6900, Saudi Arabia; [email protected] Department of Neuroscience, University of Connecticut School of Medicine, Farmington, CT 06030, USA3 King Abdullah University of Science and Technology (KAUST), Computational Bioscience Research
Center (CBRC), Thuwal 23955-6900, Saudi Arabia4 Centre de Biochimie Structurale, CNRS, INSERM, Université de Montpellier, 34090 Montpellier, France5 Department of Chemistry, Biology & Biotechnology, University of Perugia, 06121 Perugia, Italy* Correspondence: [email protected] (F.L.-C); [email protected] (C.G.)
Received: 28 March 2020; Accepted: 15 May 2020; Published: 27 May 2020�����������������
Abstract: Hyperpolarization-activated calcium channels (HACCs) are found in the plasma membraneand tonoplast of many plant cell types, where they have an important role in Ca2+-dependent signalling.The unusual gating properties of HACCs in plants, i.e., activation by membrane hyperpolarizationrather than depolarization, dictates that HACCs are normally open in the physiological hyperpolarizedresting membrane potential state (the so-called pump or P-state); thus, if not regulated, they wouldcontinuously leak Ca2+ into cells. HACCs are permeable to Ca2+, Ba2+, and Mg2+; activated byH2O2 and the plant hormone abscisic acid (ABA); and their activity in guard cells is greatly reducedby increasing amounts of free cytosolic Ca2+ ([Ca2+]Cyt), and hence closes during [Ca2+]Cyt surges.Here, we demonstrate that the presence of the commonly used Mg-ATP inside the guard cell greatlyreduces HACC activity, especially at voltages ≤ −200 mV, and that Mg2+ causes this block. Therefore,we firstly conclude that physiological cytosolic Mg2+ levels affect HACC gating and that channelopening requires either high negative voltages (≥−200 mV) or displacement of Mg2+ away from theimmediate vicinity of the channel. Secondly, based on structural comparisons with a Mg2+-sensitiveanimal inward-rectifying K+ channel, we propose that the likely candidate HACCs described hereare cyclic nucleotide gated channels (CNGCs), many of which also contain a conserved diacidicMg2+ binding motif within their pores. This conclusion is consistent with the electrophysiologicaldata. Finally, we propose that Mg2+, much like in animal cells, is an important component in Ca2+
signalling and homeostasis in plants.
Keywords: hyperpolarization-activated calcium channels; HACCs; cyclic nucleotides-activatedchannels; CNGCs; magnesium; calcium; guard cells; patch clamp; cellular homeostasis;structure modelling
1. Introduction
Ca2+ has long been recognized as an essential component in many plant cellular processes. In orderfor Ca2+ to function as an intracellular signal, temporal-, spatial-, and stimulus-specific changes in[Ca2+]Cyt need to be tightly controlled [1]. Ca2+ influx into plant cells is achieved by three maintypes of Ca2+ channels [2,3]: firstly, channels that show little or no voltage sensitivity, referred to asnon-selective calcium channels (NSCCs), primarily for being active at all voltages and for being lessselective for calcium over monovalent cations, such as K+ and Na+ [4]; secondly, channels that showhigh voltage-dependence, such as those activated by depolarization (DACCs) [5]; thirdly, others thatare activated only by membrane hyperpolarization (HACCs) [6], which are the focus of this report.
Int. J. Mol. Sci. 2020, 21, 3771; doi:10.3390/ijms21113771 www.mdpi.com/journal/ijms

Int. J. Mol. Sci. 2020, 21, 3771 2 of 19
In guard cells, Ca2+ was shown to be involved in abscisic acid (ABA) signalling and stomatalguard cell movements [7,8]. Using the patch clamp technique in both whole cell (WC) and excisedconfigurations (EC), two types of HACCs were identified in the plasma membrane (PM). One typeof HACCs is highly selective for Ca2+ (over K+ and Cl-) [9]. Its activity is enhanced by ABA [9,10],H2O2 [10], and by external Ca2+ itself, an apparently unique property of this particular plant HACC [11].Meanwhile, increasing [Ca2+]Cyt from 0.2 to 2 µM decreased the open probability (Po) of this HACC bya factor of 10 [9], implying a critical role of [Ca2+]Cyt in the feedback regulation of the channel. Theproteins responsible for this type of channel activity have not yet been identified. This is not the casefor the other type of HACCs, where a family of 20 genes is known and their translation products wereoriginally described as being mostly gated open by cyclic nucleotides, such as 3′,5′-cyclic adenosinemonophosphate (cAMP) or 3′,5′- cyclic guanosine monophate (cGMP) [12]; hence, the reason for theirname—cyclic nucleotide gated channels (CNGCs). Moreover, these plant channels, similarly to theiranimal functional homologs, poorly discriminate between divalent and monovalent cations. Indeed,when heterologously expressed in oocytes, these channels are not only permeable to Ca2+ [13] butare found to be equally permeable to monovalent cations such as K+, Rb+, Na+, Li+, and Cs+ [13,14].Similar inwardly rectifying currents (I) permeable to either Ca2+, Ba2+, and even Na+, which areactivated by cAMP [15] and cGMP [16], were also characterized in guard cells with the patch clamptechnique. The latter work demonstrated that the highly expressed CNGC5 and CNGC6 genes inguard cells are directly responsible for the recorded HACC activity. Likewise, in pollen tubes of theAsian pear tree (Pyrus pyrifolia Nakai cv. Hosui), a HACC that conducts indiscriminately either Ca2+ orK+ was shown to be activated by cAMP and downregulated by high [Ca2+]Cyt [17]. It is noteworthythat all these HACCs share one common characteristic—they are sensitive to low concentrations oflanthanides [10,15–17].
We know from previous work, particularly in animal systems, that Mg2+ ions are key regulatorsof many ion channels and receptors [18]. For instance, the inwardly rectifying K+ (Kir) channels andTRPV6, a member belonging to a subgroup of transient receptor potential (TRP) cation channels, bothshow strong Mg2+-dependent gating [19,20]. Another example of a voltage-dependent blockage byMg2+ ions is the N-methyl-D-aspartate (NMDA) receptor [21,22]; in this case extracellular Mg2+ isresponsible for this effect. By contrast, in plants, the rectification of IK,in was found not to be due toMg2+-dependent blockage [23] but rather due to an intrinsic property of the channel protein itself [24].However, ion channels localized to the tonoplast with a key role in stomatal volume regulation, suchas the slow (SV)- and the fast (FV)-activating vacuolar channels, were shown to be affected by cytosolicMg2+ ions [25–28]. Indeed, it was found that besides Ca2+, Mg2+ also promoted the activation ofSV channels, affecting their kinetics (time constants of channel activation and de-activation) andvoltage-dependent activation characteristics. At the same time, Mg2+ inhibits FV channels, thusreducing K+ leakage from the tonoplast [25]. Mg2+ was also shown to inhibit an outward NSCC,termed MgC, which was characterized in the PM of guard and subsidiary cells of Vicia faba andZea mays [29]. Another NSCC example from N2-fixing plants where Mg2+ plays a critical role isits involvement in the transport of ammonium (NH4
+) or ammonia (NH3) across the peribacteroidmembrane [30]. This is partly achieved through a passive non-selective electrogenic transport systemregulated by Ca2+. More recently, this channel was described as having “unusual” characteristics,such as an inward rectification caused by Mg2+ on the cytosolic face and a very low single channelconductance (<0.2 pS with 150 mM KCl + 10 mM CaCl2 in the pipette and 150 mM NH4
+ in the bath),which was found to be inhibited by Mg2+ from the luminal face of the symbiosome [31].
ATP is another major regulator of ion channel gating. One of the best-characterized channelsis KATP, an inward K+-rectifier (from the Kir family of genes) found mostly in cardiac and skeletalmuscles, neurons, and pancreatic cells [18]. These channels are normally closed in the presence of ATPand only open to hyperpolarize the cell membrane when cytosolic ATP levels drop. ADP added in theform of Mg-ADP can restore the activity of KATP pre-treated with ATP [32,33]. In guard cells, Mg-ATPis required for blue light-activated outward currents [34]. Indeed, it was found that 1 to 2 mM Mg-ATP,

Int. J. Mol. Sci. 2020, 21, 3771 3 of 19
as well as other intracellular substrates, are required to fully activate a plasma membrane electrogenicion pump capable of hyperpolarizing the membrane to around −140 mV, a potential well beyond theactivation threshold for IK,in [35].
Given the importance of calcium channels in plant cellular signalling, including in guard cellaperture regulation, we address the question of whether internal Mg2+ can affect the activity HACCsin guard cell protoplasts (GCPs) and show examples of Gd3+-sensitive Ba2+ currents (IBa) activatedby hyperpolarization with and without Mg2+ in the patch pipet. We also assess important propertiesof IBa in the absence of Mg2+, such as the permeability and sensitivity to some relevant inorganiccompounds and physiological effectors, including abscisic acid (ABA) and cAMP.
2. Materials and Methods
2.1. Protoplast Isolation
Vicia faba L. cv (Bunyan) Bunyan Exhibition seeds were grown on vermiculite under conditionsdescribed previously [36]. Arabidopsis thaliana (Columbia) seeds were grown on peat pellets (jiffy, Oslo)in a controlled environment growth chamber (Percival, CLF plant climatic, Wertingen) at 22 ◦C on a8/16-h light/dark cycle. Guard cell protoplasts (GCPs) were isolated from either week 3 or 4 in V. fabaor from week 5 or 6 in Arabidopsis thaliana plants. GCPs were isolated from abaxial epidermal strips asdescribed previously [37]. Briefly, epidermal strips were floated on medium containing 1.8–2.5% (w/v)Cellulase Onozuka RS (Yacult Honsha, Tokyo, Japan), 1.7–2% (w/v) Cellulysin (Calbiochem, BehringDiagnostics, La Jolla, CA, USA), 0.026% (w/v) Pectolyase Y-23, 0.26% (w/v) BSA, and 1 mM CaCl2(pH 5.6), osmolality adjusted with sorbitol to 360 mOsm.kg−1. After 90–120 min incubation in the darkat 28 ◦C with gentle shaking, released protoplasts were passed through a 30 µm mesh and kept on icefor 2 to 3 min before centrifugation (100 g for 4 min at room temperature). The pellet consisting of GCPswas re-suspended and kept on ice in 1 or 2 mL of fresh medium containing 0.42 M mannitol, 10 mM2-(N-morpholino) ethanesulfonic acid (Mes), 200 µM CaCl2, 2.5 mM KOH (pH 5.55 and osmolality at466 mOsm.kg−1). Unless stated otherwise, all chemicals were from Sigma (Sigma-Aldrich Co., St Louis,MO, USA).
2.2. Solutions
Protoplasts were placed in a 0.5 mL chamber, left to settle down, and then perfused continuouslyat a flow rate of ≈0.5–1 mL/min. To record IBa currents through HACC, we used barium-containingsolutions. The bath medium contained (in mM): 100 BaCl2, 10 Mes (pH 5.5 with Tris base); the pipettecontained (in mM): 100 BaCl2, 4 EGTA, 10 Hepes (pH 7.5 with Tris). For experiments where IK,in andIBa measurements were made on the same GCPs, a different bath and internal solutions as follows.Bath (in mM): 30 KCl, 10 Mes (pH 5.5 with Tris base) to measure IK,in, which was replaced by 100BaCl2, 10 Mes (pH 5.5 with Tris base) to measure IBa. Internal solution (in mM): 1 BaCl2, 18 KCl,4 EGTA, 10 Hepes (pH 7.5 with Tris base). Mg-ATP, MgCl2, or K2-ATP were added as specified inthe figures. Osmolality was adjusted with sorbitol to 310 mOsm.kg−1. For classic solutions used tomeasure IK,in refer to [38]. ABA was added externally. All chemicals were from Sigma Chemical, Poole,Dorset, UK. The membrane permeable cAMP analog Bt2cAMP was solubilized in deionized water andstored in aliquots of 50–100 µL at a concentration of 0.1 M. Bt2cAMP was diluted to the final desiredconcentration just a few minutes before its use.
2.3. Current–Voltage Recording and Analysis
Patch pipettes (5–10) were pulled from Kimax-51 glass capillaries (Kimble 34500; Kimble,Owens-Illinois) using a two-stage puller (Narishige PP-83, Japan). Experiments were performed atroom temperature (20 to 22 ◦C) using standard whole-cell patch clamp techniques, with an Axopatch200B integrating patch clamp amplifier (Axon Instruments, Inc., Union City, CA, USA). Voltagecommands and simultaneous signal recordings and analyses were assessed by a microcomputer

Int. J. Mol. Sci. 2020, 21, 3771 4 of 19
connected to the amplifier via a multipurpose input–output device (Digidata 1320A) using pClampsoftware (versions 8.0 and 10; Axon Instruments, Inc.). After giga ohm seals were formed, the whole-cellconfiguration was then achieved by gentle suction and the membrane was immediately clamped to aholding voltage (hv) of −36 mV. GCPs were continuously perfused throughout the experiment andcurrent recordings began only after at least 5–10 min from going into whole-cell mode to allow forintracellular equilibrium between the cytoplasm and patch pipet solution. All current traces shownwere low-pass filtered at 2 kHz before analog-to-digital conversion and were uncorrected for leakagecurrent or capacitive transients. Membrane potentials were corrected for liquid junction potential asdescribed in [39]. Ionic activities were calculated using GEOCHEM-EZ [40]. Current–voltage (I-V)relationships for IBa and IK,in were plotted as steady-state currents vs. test potentials when using thesquare pulse stimulations or utilizing the “trace vs. trace” feature of Clampfit analysis when usingvoltage ramps. Unless otherwise stated, every experiment reported here was repeated a minimum ofthree times and data were graphed as mean ± SEM.
2.4. Protein Structural Modelling
Models were built using Swiss-Model [41] and structures were visualized with PyMOL (PyMOLMolecular Graphics System, Version 1.5.0.4 Schrödinger, LLC). Homology modelling was supportedby PSIPRED [42] and multisequence alignments were produced with MUSCLE [43]. Alignments andadditional information of the models are provided in the respective legends.
3. Results
3.1. Removal of Intracellular Mg-ATP Unveils a Larger Instantaneously-Activated, Inwardly-Directed,and Gd3+-Sensitive Ba2+ Current
We recorded Ba2+ currents (IBa) in guard cell protoplasts in either the presence or absence ofMg-ATP (Figure 1A). At the end of the trials we also tested for IBa sensitivity to Gd3+, a potent blockerof IBa (Figure 1B). The recorded currents were generated in response to a square pulse protocol from+64 mV to −256 mV in increments of −20 mV, with the holding voltage (hV) set to −36 mV. In thepresence of Mg-ATP (Figure 1A; left panel), a small instantaneous inward-rectifying current of ≈−20 pAstarted to activate at around −200 mV, reaching a maximum of −60 pA at −256 mV. As expected, thiscurrent was sensitive to the addition of extracellular Gd3+ (see Figure 1B; left panel). Meanwhile, in theabsence of Mg-ATP (Figure 1A; right panel), much larger (≥8-fold) instantaneous rectifying currentswere recorded, which happened to also be Gd3+-sensitive (Figure 1B; left traces). The current–voltage(I-V) relationships in the presence (•; n = 3) or absence (#; n = 7) of Mg-ATP are represented inFigure 1C (left panel). We also plotted the effect of Gd3+ on the I-V relationships in the presence (�) orabsence (�) of Mg-ATP (Figure 1C; right panel).

Int. J. Mol. Sci. 2020, 21, 3771 5 of 19Int. J. Mol. Sci. 2020, 21, x 5 of 19
Figure 1. Removal of intracellular Mg-ATP unveils a larger instantaneously-activated, inwardly-directed, and Gd3+-sensitive Ba2+ current. (A) Typical examples of current traces in whole-cell mode obtained from two separate V. faba guard cells with the pipette solution either containing (left traces) or lacking (right traces) Mg-ATP (1 mM). The pulse protocol mostly used throughout this study consisted of 0.6 s long square voltage pulses ranging from +64 to −256 mV in −20 mV increments; the holding potential hv was set to −36 mV. In order to preserve the quality of the “giga” seals, guard cell protoplasts (GCPs) with no Mg-ATP in the pipette were not subjected to higher voltages beyond −196 mV. (B) Current (I) traces of IBa obtained from the same cells and in the same conditions as described in (A), except for the external solution containing Gd3+ (20 mM, left; or 100 mM, right) (C) Current–voltage relationships (I-V): left panel, superimposed I-Vs of IBa in the absence (○; n = 7) or presence (●: n = 3) of Mg-ATP; right panel, I-V relations in the presence of Gd3+ (■: +Mg-ATP; □: −Mg-ATP). (D) The reversal potential (Erev) is close to the reversal potential for Ba2+ (EBa). Typical current trace recorded in the absence of Mg-ATP (below) in response to a voltage ramp which consists of activating IBa using a hyperpolarization square pulse to −156 mV and immediately followed by a continuous depolarizing ramp to +64 mV (the voltage protocol is depicted above), with a slope of 0.7 V s−1. Scale bars are shown below the current traces and to the left of the voltage protocol.
These results highlight that when omitting Mg-ATP from the intracellular medium, a larger Gd3+-sensitive, inwardly rectifying Ba2+ current is unveiled, which activates at significantly less-
-20 20 40 60
-5
5
V (mV)
I (pA)
-256 mV
50 pA0.1 s
-196 mV
A
B
0 Mg-ATP+ Mg-ATP
-200 -100
-200
-100
V (mV) I (pA
)-200 -100
-200
-100
V (mV)
I (pA)
-256 mV -196 mV
C
-50
0
(pA
)
0.4 s
0.2 V
EBaECl
D
- Gd3+
+ Gd3+
+ Mg-ATP
0 Mg-ATP
+Mg-ATP 0 Mg-ATP
- Gd3+ + Gd3+
Figure 1. Removal of intracellular Mg-ATP unveils a larger instantaneously-activated,inwardly-directed, and Gd3+-sensitive Ba2+ current. (A) Typical examples of current traces in whole-cellmode obtained from two separate V. faba guard cells with the pipette solution either containing (lefttraces) or lacking (right traces) Mg-ATP (1 mM). The pulse protocol mostly used throughout this studyconsisted of 0.6 s long square voltage pulses ranging from +64 to −256 mV in −20 mV increments;the holding potential hv was set to −36 mV. In order to preserve the quality of the “giga” seals, guardcell protoplasts (GCPs) with no Mg-ATP in the pipette were not subjected to higher voltages beyond−196 mV. (B) Current (I) traces of IBa obtained from the same cells and in the same conditions asdescribed in (A), except for the external solution containing Gd3+ (20 mM, left; or 100 mM, right) (C)Current–voltage relationships (I-V): left panel, superimposed I-Vs of IBa in the absence (#; n = 7) orpresence (•: n = 3) of Mg-ATP; right panel, I-V relations in the presence of Gd3+ (�: +Mg-ATP; �:−Mg-ATP). (D) The reversal potential (Erev) is close to the reversal potential for Ba2+ (EBa). Typicalcurrent trace recorded in the absence of Mg-ATP (below) in response to a voltage ramp which consistsof activating IBa using a hyperpolarization square pulse to −156 mV and immediately followed by acontinuous depolarizing ramp to +64 mV (the voltage protocol is depicted above), with a slope of0.7 V s−1. Scale bars are shown below the current traces and to the left of the voltage protocol.
These results highlight that when omitting Mg-ATP from the intracellular medium, a largerGd3+-sensitive, inwardly rectifying Ba2+ current is unveiled, which activates at significantlyless-negative voltages (see the shift to the right of ≥−100 mV in the I-V plot). Furthermore, thecurrents recorded in 0 Mg-ATP seem to reverse near the calculated Nernst equilibrium potential for

Int. J. Mol. Sci. 2020, 21, 3771 6 of 19
Ba2+ (EBa ≈ +28 mV), and are far removed from ECl (−54 mV). Likewise, using fast depolarizationramps (0.07 V/s) after activating the current IBa with a square pulse to −156 mV (see voltage protocoland current trajectories in Figure 1D; left), a reversal potential of +17 mV was measured, again close toEBa rather than ECl (see Figure 1D, zoomed I-V plot). This is also indicative of the higher permeabilityof this conductance to Ba2+ as compared to Cl−.
To test whether changing external Ba2+ concentration will affect current magnitude, as well asthe I-V relationship, GCPs were patched in whole cell mode using the Mg-ATP-free internal solution(Figure 2).
Int. J. Mol. Sci. 2020, 21, x 6 of 19
negative voltages (see the shift to the right of ≥−100 mV in the I–V plot). Furthermore, the currents recorded in 0 Mg-ATP seem to reverse near the calculated Nernst equilibrium potential for Ba2+ (EBa
≈ +28 mV), and are far removed from ECl (−54 mV). Likewise, using fast depolarization ramps (0.07 V/s) after activating the current IBa with a square pulse to −156 mV (see voltage protocol and current trajectories in Figure 1D; left), a reversal potential of +17 mV was measured, again close to EBa rather than ECl (see Figure 1D, zoomed I-V plot). This is also indicative of the higher permeability of this conductance to Ba2+ as compared to Cl−.
To test whether changing external Ba2+ concentration will affect current magnitude, as well as the I-V relationship, GCPs were patched in whole cell mode using the Mg-ATP-free internal solution (Figure 2).
Figure 2. Decreasing [Ba2+]o from 100 to 30 mM not only decreases the hyperpolarization-activated IBa but also shifts its apparent reversal potential (Erev) (~−14 mV). (A,B) Typical current traces of hyperpolarization-activated IBa recorded from the same V. faba guard cell in either 100 (A) or 30 mM (B) [Ba2+]o. Scale bars are shown below the current traces. (C) Corresponding I-V plots of IBa in 100 and 30 mM [Ba2+]o taken from the current traces shown in (A) and (B). Inset: Magnified I-V plot from the area shown as a dashed box in (C). The inset shows the amount (in mV) and the direction (arrow) of the shift in the apparent Erev when the bath perfusion was switched from 100 to 30 mM Ba2+.
Once again, the current magnitudes recorded in the absence of Mg-ATP are substantial. For instance, at −196 mV, a −280 pA current is measured in 100 mM [Ba2+]out (Figure 2A), while in 30 mM [Ba2+]out (Figure 2B) the same voltage gives rise to a current value of −220 pA. All current magnitudes at any given voltage are decreased when switching to lower [Ba2+] in the bath. The corresponding I-V plots appear as shifting to negative values (Figure 2C) when switching from 100 to 30 mM Ba2+, which is accompanied by a negative shift in the apparent reversal potential (Erev) values. Indeed, when zooming in (Figure 2C, inset), the apparent Erev shows a negative shift of ≈ −14 mV as a result of this Ba2+ concentration change. Furthermore, the apparent Erev in 30 mM [Ba2+]out (≈ +8 mV) is still closer to the calculated EBa rather than ECl, the values for which in this case are +17.7 and −25.7 mV,
-180 -120 -60 60
-300
-150
V (mV)
I (pA)
Ba 100 Ba 30
C
A100 BaCl2
0.1 s
0.1 nA
30 BaCl2B
-20 -10 10 20 30 40 50
-6
6
V (mV)
I (p
A)
Shift of apparent Erevfor Barium (-14mV)
Figure 2. Decreasing [Ba2+]o from 100 to 30 mM not only decreases the hyperpolarization-activatedIBa but also shifts its apparent reversal potential (Erev) (∼−14 mV). (A,B) Typical current traces ofhyperpolarization-activated IBa recorded from the same V. faba guard cell in either 100 (A) or 30 mM (B)[Ba2+]o. Scale bars are shown below the current traces. (C) Corresponding I-V plots of IBa in 100 and30 mM [Ba2+]o taken from the current traces shown in (A) and (B). Inset: Magnified I-V plot from thearea shown as a dashed box in (C). The inset shows the amount (in mV) and the direction (arrow) ofthe shift in the apparent Erev when the bath perfusion was switched from 100 to 30 mM Ba2+.
Once again, the current magnitudes recorded in the absence of Mg-ATP are substantial. Forinstance, at −196 mV, a −280 pA current is measured in 100 mM [Ba2+]out (Figure 2A), while in 30 mM[Ba2+]out (Figure 2B) the same voltage gives rise to a current value of −220 pA. All current magnitudesat any given voltage are decreased when switching to lower [Ba2+] in the bath. The correspondingI-V plots appear as shifting to negative values (Figure 2C) when switching from 100 to 30 mM Ba2+,which is accompanied by a negative shift in the apparent reversal potential (Erev) values. Indeed,when zooming in (Figure 2C, inset), the apparent Erev shows a negative shift of ≈−14 mV as a result ofthis Ba2+ concentration change. Furthermore, the apparent Erev in 30 mM [Ba2+]out (≈+8 mV) is stillcloser to the calculated EBa rather than ECl, the values for which in this case are +17.7 and −25.7 mV,respectively. This experiment was repeated with 100, 30, and 10 mM BaCl2 in the bath, with thesame qualitative effects observed, i.e., decrease of current amplitude when decreasing the [Ba2+]o andnegative shift in the apparent Erev.
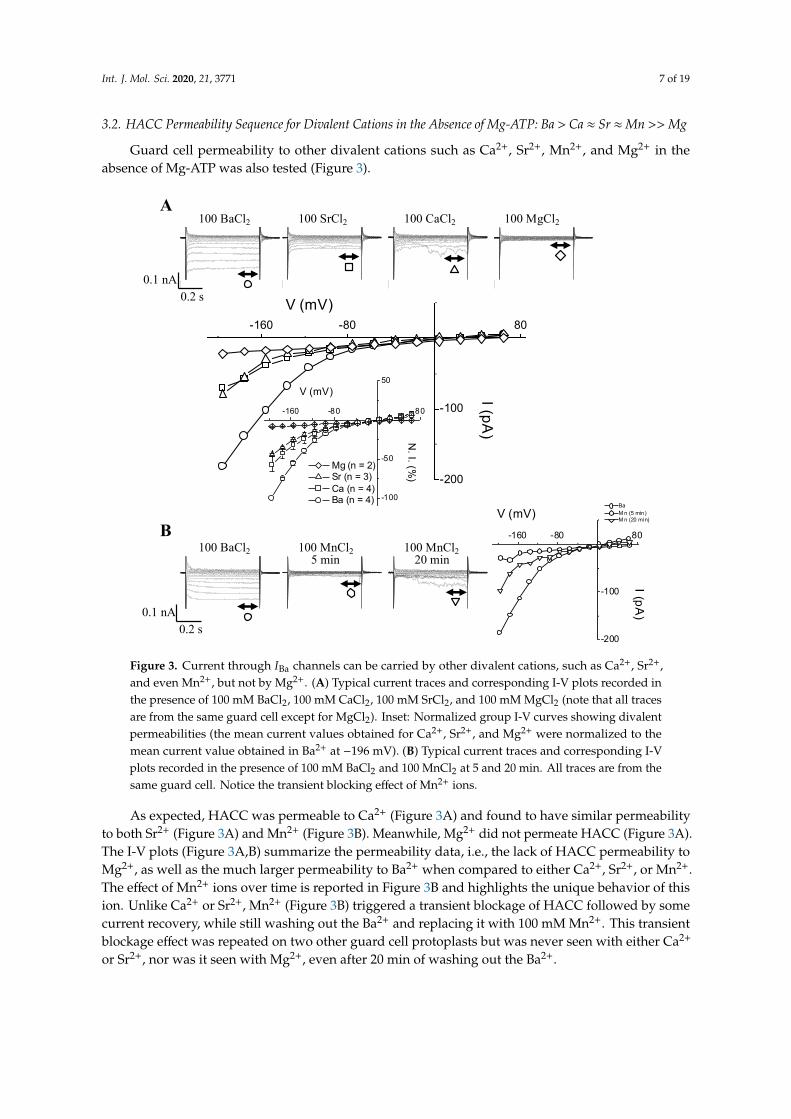
Int. J. Mol. Sci. 2020, 21, 3771 7 of 19
3.2. HACC Permeability Sequence for Divalent Cations in the Absence of Mg-ATP: Ba > Ca ≈ Sr ≈Mn >> Mg
Guard cell permeability to other divalent cations such as Ca2+, Sr2+, Mn2+, and Mg2+ in theabsence of Mg-ATP was also tested (Figure 3).
Int. J. Mol. Sci. 2020, 21, x 7 of 19
respectively. This experiment was repeated with 100, 30, and 10 mM BaCl2 in the bath, with the same qualitative effects observed, i.e., decrease of current amplitude when decreasing the [Ba2+]o and negative shift in the apparent Erev.
3.2. HACC Permeability Sequence for Divalent Cations in the Absence of Mg-ATP: Ba > Ca ≈ Sr ≈ Mn >> Mg
Guard cell permeability to other divalent cations such as Ca2+, Sr2+, Mn2+, and Mg2+ in the absence of Mg-ATP was also tested (Figure 3).
Figure 3. Current through IBa channels can be carried by other divalent cations, such as Ca2+, Sr2+, and even Mn2+, but not by Mg2+. (A) Typical current traces and corresponding I-V plots recorded in the presence of 100 mM BaCl2, 100 mM CaCl2, 100 mM SrCl2, and 100 mM MgCl2 (note that all traces are from the same guard cell except for MgCl2). Inset: Normalized group I-V curves showing divalent permeabilities (the mean current values obtained for Ca2+, Sr2+, and Mg2+ were normalized to the mean current value obtained in Ba2+ at −196 mV). (B) Typical current traces and corresponding I-V plots recorded in the presence of 100 mM BaCl2 and 100 MnCl2 at 5 and 20 minutes. All traces are from the same guard cell. Notice the transient blocking effect of Mn2+ ions.
As expected, HACC was permeable to Ca2+ (Figure 3A) and found to have similar permeability to both Sr2+ (Figure 3A) and Mn2+ (Figure 3B). Meanwhile, Mg2+ did not permeate HACC (Figure 3A). The I-V plots (Figure 3A,B) summarize the permeability data, i.e., the lack of HACC permeability to Mg2+, as well as the much larger permeability to Ba2+ when compared to either Ca2+, Sr2+, or Mn2+. The effect of Mn2+ ions over time is reported in Figure 3B and highlights the unique behavior of this ion. Unlike Ca2+ or Sr2+, Mn2+ (Figure 3B) triggered a transient blockage of HACC followed by some current recovery, while still washing out the Ba2+ and replacing it with 100 mM Mn2+. This transient blockage
-160 -80 80
-200
-100
Ba M n (5 min ) M n (20 min)
V (mV)
I (pA)
100 MgCl2100 BaCl2 100 CaCl2
0.2 s0.1 nA
A100 SrCl2
-160 -80 80
-200
-100
V (mV) I (pA
)
B100 BaCl2 100 MnCl2100 MnCl2
5 min 20 min
0.2 s0.1 nA
-160 -80 80
-100
-50
50
Mg (n = 2) Sr (n = 3) Ca (n = 4) Ba (n = 4)
V (mV) N
. I. (%)
Figure 3. Current through IBa channels can be carried by other divalent cations, such as Ca2+, Sr2+,and even Mn2+, but not by Mg2+. (A) Typical current traces and corresponding I-V plots recorded inthe presence of 100 mM BaCl2, 100 mM CaCl2, 100 mM SrCl2, and 100 mM MgCl2 (note that all tracesare from the same guard cell except for MgCl2). Inset: Normalized group I-V curves showing divalentpermeabilities (the mean current values obtained for Ca2+, Sr2+, and Mg2+ were normalized to themean current value obtained in Ba2+ at −196 mV). (B) Typical current traces and corresponding I-Vplots recorded in the presence of 100 mM BaCl2 and 100 MnCl2 at 5 and 20 min. All traces are from thesame guard cell. Notice the transient blocking effect of Mn2+ ions.
As expected, HACC was permeable to Ca2+ (Figure 3A) and found to have similar permeabilityto both Sr2+ (Figure 3A) and Mn2+ (Figure 3B). Meanwhile, Mg2+ did not permeate HACC (Figure 3A).The I-V plots (Figure 3A,B) summarize the permeability data, i.e., the lack of HACC permeability toMg2+, as well as the much larger permeability to Ba2+ when compared to either Ca2+, Sr2+, or Mn2+.The effect of Mn2+ ions over time is reported in Figure 3B and highlights the unique behavior of thision. Unlike Ca2+ or Sr2+, Mn2+ (Figure 3B) triggered a transient blockage of HACC followed by somecurrent recovery, while still washing out the Ba2+ and replacing it with 100 mM Mn2+. This transientblockage effect was repeated on two other guard cell protoplasts but was never seen with either Ca2+
or Sr2+, nor was it seen with Mg2+, even after 20 min of washing out the Ba2+.

Int. J. Mol. Sci. 2020, 21, 3771 8 of 19
3.3. HACC Permeability Sequence for Monovalent Cations in the Absence of Mg-ATP: K ≈ Na > Ba > Cs>> TEA
We observed that this Gd3+-sensitive HACC is also permeable to some physiologically relevantmonovalent cations, such as K+ (Figure S1A), Na+ (Figure S1B), and Cs+ (Figure S1C), but nottetraethylammonium (TEA+) (Figure S1D). These data indicate that the Gd3+-sensitive currentcharacterized in this work does not select for small mono or divalent cations (except for the case ofMg2+ and the bigger cation TEA+). Chloride also does not seem to permeate through this HACC.Indeed, when 100 mM Cl- was added at the same time as TEA+ (see Figure S1D), no current could bedetected, indicating that Cl- is as impermeable as TEA+. Qualitatively, the same effect was seen in allpatched GCPs (n = 5 for K+ and Na+; n = 2 for Cs+ and TEA+).
3.4. Effect of Blockers of IBa in the Absence of Mg-ATP and Comparison with the Effect on IK,in
In order to further characterize this HACC, which was readily unmasked when Mg-ATP wasomitted from the patch pipet, the effect of some classical blockers such as the lanthanides (La3+ andGd3+), Mn2+, Cs+, and TEA+ were tested on IBa, as well as on IK,in, (the other major conductance thatactivates upon hyperpolarization in guard cells) (Figure 4).
Int. J. Mol. Sci. 2020, 21, x 8 of 19
effect was repeated on two other guard cell protoplasts but was never seen with either Ca2+ or Sr2+, nor was it seen with Mg2+, even after 20 minutes of washing out the Ba2+.
3.3. HACC Permeability Sequence for Monovalent Cations in the Absence of Mg-ATP: K ≈ Na > Ba > Cs >> TEA.
We observed that this Gd3+-sensitive HACC is also permeable to some physiologically relevant monovalent cations, such as K+ (Figure S1A), Na+ (Figure S1B), and Cs+ (Figure S1C), but not tetraethylammonium (TEA+) (Figure S1D). These data indicate that the Gd3+-sensitive current characterized in this work does not select for small mono or divalent cations (except for the case of Mg2+ and the bigger cation TEA+). Chloride also does not seem to permeate through this HACC. Indeed, when 100 mM Cl- was added at the same time as TEA+ (see Figure S1D), no current could be detected, indicating that Cl- is as impermeable as TEA+. Qualitatively, the same effect was seen in all patched GCPs (n = 5 for K+ and Na+; n = 2 for Cs+ and TEA+).
3.4. Effect of Blockers of IBa in the Absence of Mg-ATP and Comparison with the Effect on IK,in.
In order to further characterize this HACC, which was readily unmasked when Mg-ATP was omitted from the patch pipet, the effect of some classical blockers such as the lanthanides (La3+ and Gd3+), Mn2+, Cs+, and TEA+ were tested on IBa, as well as on IK,in, (the other major conductance that activates upon hyperpolarization in guard cells) (Figure 4).
Figure 4. Differential effects of some known blockers on the two main PM conductances activated by hyperpolarization in guard cells, namely IBa and IK,in. (A) I-V plots showing the effect of Gd3+ (0.1 mM) on IBa (left panel) and IK,in (right panel). (B) I-V plots showing the effect of 1 mM Cs+ on IBa (left panel) compared to the effect of 0.1 mM Cs+ on IK,in (right panel). (C) I-V plot showing the effect of Mn2+ on IBa recorded from the same GCP using 2 and 10 mM in the bath. (D) IBa-V plots generated from current recordings using hyperpolarizing ramps (+64 to −196 mV; 0.7 V.s−1; hv = −36 mV), showing the effect of 10 mM TEA+ added to the bath.
One of the most conspicuous effects lies in the potent effect of Gd3+ in blocking IBa (Figure 4A), even when used at relatively low concentrations (20 to 100 µM), while the same concentrations of
-160 -80
-400
-200
V (mV)
I (pA)
-160 -80
-300
-150
V (mV)
I (pA)
-160 -80 80
-100
V (mV)
I (pA)
-160 -80 80
-100
V (mV)
I (pA)
+Gd3+
0.1 mM
+Cs+
0.1 mM
-Cs+
+Cs+
1 mM
+Gd3+
0.1 mM
-Cs+
-Gd3+
IBa
-Gd3+
B
-160 -80 80
-100
V (mV)
I (pA)
A
CV (mV)
I (pA)
-100
-50
-160 -80+80
D
IK,in
IBa IK,in
IBa IBa
-TEA+
+TEA+
10 mM-Mn2+
10 mM2 mM
+Mn2+
Figure 4. Differential effects of some known blockers on the two main PM conductances activated byhyperpolarization in guard cells, namely IBa and IK,in. (A) I-V plots showing the effect of Gd3+ (0.1 mM)on IBa (left panel) and IK,in (right panel). (B) I-V plots showing the effect of 1 mM Cs+ on IBa (left panel)compared to the effect of 0.1 mM Cs+ on IK,in (right panel). (C) I-V plot showing the effect of Mn2+ onIBa recorded from the same GCP using 2 and 10 mM in the bath. (D) IBa-V plots generated from currentrecordings using hyperpolarizing ramps (+64 to −196 mV; 0.7 V.s−1; hv = −36 mV), showing the effectof 10 mM TEA+ added to the bath.
One of the most conspicuous effects lies in the potent effect of Gd3+ in blocking IBa (Figure 4A),even when used at relatively low concentrations (20 to 100 µM), while the same concentrations ofGd3+ had no effect on IK,in (Figure 4A). An even higher concentration of Gd3+ (500 µM) did not affect
Int. J. Mol. Sci. 2020, 21, 3771 9 of 19
IK,in (data not shown). La3+ also blocked IBa measured in 0 Mg-ATP, but we found that much higherconcentrations of La3+ (0.2 to 0.5 mM) are needed to achieve the same block as compared to Gd3+ (datanot shown). Experimenting with cesium, a blocker of IK,in, hardly any effect on IBa was registered(Figure 4B). Even though Cs+ was used at concentrations up to 1 mM, it had no or only a small effecton IBa, while one-tenth of this amount (0.1 mM) is sufficient to block a large proportion (≈ 80% ormore) of IK,in (Figure 4B). Furthermore, Mn2+ used at 2 mM inhibited HACC by ≈37% (at V = −196mV) when the charge carrier (in this case Ba2+) was still present in the bath (Figure 4C). IncreasingMn2+ concentration to 10 mM shows that Mn2+ is not an efficient blocker of IBa as compared to Gd3+
or La3+, and 10 mM Mn2+ only causes an extra 20% IBa inhibition (see IV plot in Figure 4C). Finally, 10mM tetraethylammonium chloride (TEA), a concentration that was shown to block 70% to 80% of IK,in
in intact guard cells [44], had no effect whatsoever on IBa (Figure 4D) measured in 0 Mg-ATP.
3.5. Rapid Enhancement of IBa by ABA
Given that in V. faba guard cells a HACC was implicated downstream of ABA in stomatalmovements, we tested whether ABA affects IBa activated in the absence of internal Mg-ATP. Wepatch-clamped guard cells to measure IBa currents under baseline conditions, i.e., zero Mg-ATP insideand no added ABA outside (Figure 5).
Int. J. Mol. Sci. 2020, 21, x 9 of 19
Gd3+ had no effect on IK,in (Figure 4A). An even higher concentration of Gd3+ (500 µM) did not affect IK,in (data not shown). La3+ also blocked IBa measured in 0 Mg-ATP, but we found that much higher concentrations of La3+ (0.2 to 0.5 mM) are needed to achieve the same block as compared to Gd3+ (data not shown). Experimenting with cesium, a blocker of IK,in, hardly any effect on IBa was registered (Figure 4B). Even though Cs+ was used at concentrations up to 1 mM, it had no or only a small effect on IBa, while one-tenth of this amount (0.1 mM) is sufficient to block a large proportion (≈ 80% or more) of IK,in (Figure 4B). Furthermore, Mn2+ used at 2 mM inhibited HACC by ≈ 37% (at V = −196 mV) when the charge carrier (in this case Ba2+) was still present in the bath (Figure 4C). Increasing Mn2+ concentration to 10 mM shows that Mn2+ is not an efficient blocker of IBa as compared to Gd3+ or La3+, and 10 mM Mn2+ only causes an extra 20% IBa inhibition (see IV plot in Figure 4C). Finally, 10 mM tetraethylammonium chloride (TEA), a concentration that was shown to block 70% to 80% of IK,in in intact guard cells [44], had no effect whatsoever on IBa (Figure 4D) measured in 0 Mg-ATP.
3.5. Rapid Enhancement of IBa by ABA
Given that in V. faba guard cells a HACC was implicated downstream of ABA in stomatal movements, we tested whether ABA affects IBa activated in the absence of internal Mg-ATP. We patch-clamped guard cells to meas
Figure 5. Rapid enhancement of IBa by abscisic acid (ABA). (A) The IBa currents in the absence of Mg-ATP in the patch pipet recorded from the same guard cell in response to hyperpolarizing voltages (from +64 to −196 mV; in −20 mV increments) before (○) and 10 minutes after (●) bath application of ABA (20 µM). (B) I–V plots of the effect of ABA, showing the enhancing effect of ABA with time (control: -ABA; 5 and 10 minutes after continuous bath perfusion with ABA). We also superimposed the measurements generated by voltage ramps for the control, 5 and 10 min ABA. Inset: Superimposed I-V plots showing the average effect of ABA on IBa (0 Mg-ATP). Data are current average measurements (± SEM) from different experiments (n = 3) before () and approximately 10 minutes after () bath perfusion with ABA (Student’s test; * p ≤ 0.05, ** p ≤ 0.01, ns p > 0.05).
After about 10 minutes, the time usually necessary to reach steady-state conditions, we switched the perfusion solution to the one containing 20 µM ABA. A rapid and pronounced increase of IBa currents is seen at all voltages between −100 and −200 mV (≥ 1.3- to 1.5-fold) after only 5 minutes of ABA treatment (Figure 5B), and a near doubling of the size of the IBa currents occurs at 10 minutes
-150 -100 -50 50
-150
-100
-50
50 V (mV)
I (pA)
- ABA
+ ABA5 min
+ ABA10 min
- ABA + ABA (10 min)
50 pA100 ms
A
B
-200 -150 -100 -50 50
-100
-50
V (mV)
I (pA)
- ABA
+ ABA
Figure 5. Rapid enhancement of IBa by abscisic acid (ABA). (A) The IBa currents in the absence ofMg-ATP in the patch pipet recorded from the same guard cell in response to hyperpolarizing voltages(from +64 to −196 mV; in −20 mV increments) before (#) and 10 min after (•) bath application ofABA (20 µM). (B) I-V plots of the effect of ABA, showing the enhancing effect of ABA with time(control: -ABA; 5 and 10 min after continuous bath perfusion with ABA). We also superimposed themeasurements generated by voltage ramps for the control, 5 and 10 min ABA. Inset: Superimposed I-Vplots showing the average effect of ABA on IBa (0 Mg-ATP). Data are current average measurements(±SEM) from different experiments (n = 3) before (#) and approximately 10 min after (•) bath perfusionwith ABA (Student’s test; * p ≤ 0.05, ** p ≤ 0.01, ns p > 0.05).
After about 10 min, the time usually necessary to reach steady-state conditions, we switched theperfusion solution to the one containing 20 µM ABA. A rapid and pronounced increase of IBa currentsis seen at all voltages between −100 and −200 mV (≥1.3- to 1.5-fold) after only 5 min of ABA treatment

Int. J. Mol. Sci. 2020, 21, 3771 10 of 19
(Figure 5B), and a near doubling of the size of the IBa currents occurs at 10 min (Figure 5A,B). Theenhancement of IBa in response to ABA, especially at 10 min, spans from −60 to −200 mV, and alsoappears to shift the activation threshold of IBa (Figure 5B) to the right. This suggests that ABA not onlyenhances calcium entry through HACCs but can also mobilize calcium entry at less negative voltages.
3.6. Characterization of the Effect of Mg2+ on IBa and IK,in
To answer whether internal Mg2+ alone causes the block of IBa when we add Mg-ATP, GCPs werepatched either without Mg-ATP or without ATP, but with added Mg2+ (added as MgCl2) (Figure 6).
Int. J. Mol. Sci. 2020, 21, x 10 of 19
(Figure 5A,B). The enhancement of IBa in response to ABA, especially at 10 minutes, spans from -60 to −200 mV, and also appears to shift the activation threshold of IBa (Figure 5B) to the right. This suggests that ABA not only enhances calcium entry through HACCs but can also mobilize calcium entry at less negative voltages.
3.6. Characterization of the Effect of Mg2+ on IBa and IK,in
To answer whether internal Mg2+ alone causes the block of IBa when we add Mg-ATP, GCPs were patched either without Mg-ATP or without ATP, but with added Mg2+ (added as MgCl2) (Figure 6).
Figure 6. Internal Mg2+ is responsible for IBa inhibition. (A) Currents (left panel) and corresponding I-V plots (right panel) from the same guard cell recorded in response to hyperpolarizing voltage steps (from +64 to −196 mV; in −20 mV increments; hv = −36 mV) and in the absence of Mg-ATP using either K+ (30 mM) or Ba2+ (100 mM) as charge carriers. Notice the larger IBa compared to IK,in. (B) Current (left panel) and corresponding I-V relationships (right panel) recorded in the same external conditions as in (A), but this time using only Mg2+ ions in the patch solution (1 mM added as MgCl2) and no added ATP. In these conditions, the predominant current activated by hyperpolarization is IK,in, not IBa.
Furthermore, and as a control for the “ion transport functionality” of the patched GCPs, we used conditions that allow measurements of not just IBa, but also to record IK,in. The experiments were started in conditions allowing to probe for IK,in with KCl (30 mM) in the bath, which was then followed by replacing the KCl with a solution containing BaCl2 (100 mM). This was done first in the absence of both intracellular Mg2+ and ATP (Figure 6A), and then repeated on another batch of guard cells with internal medium containing 1 mM MgCl2 but no ATP (Figure 6B). Firstly, in zero Mg-ATP, the only current that activated in response to hyperpolarization was IBa (Figure 6A). IK,in could not be activated. Secondly, when only Mg2+ was included in the pipet solution (no ATP added), IK,in could then be activated in 30 mM K+. However, when switching the bath from K+- to Ba2+-containing media, IBa currents vanished, indicating that including only Mg2+ in the patch pipet can cause the blockage of IBa at voltages where it is normally activated in 0 Mg-ATP.
3.7. Can cAMP Activate HACCs in V. faba Guard Cells Despite the Presence of Intracellular Mg2+?
We know from our own previous work [15] that cAMP activated a Gd3+-sensitive HACC in guard cells while recording in Mg2+- and ATP-free media (Figure 7A). However, since we had shown that this conductance discriminates poorly between divalent and monovalent cations, a hallmark
-100 100
-400
-200
V (mV)
I (pA)
-200 -100 100
-150
-75
V (mV)
I (pA)
0 Mg-ATP
30 KCl 100 BaCl2
+ Mg (No ATP)
30 KCl 100 BaCl2
50 pA100 ms
30 K
100 Ba
A
B
0.2 nA0.2 s
30 K
100 Ba
Figure 6. Internal Mg2+ is responsible for IBa inhibition. (A) Currents (left panel) and correspondingI-V plots (right panel) from the same guard cell recorded in response to hyperpolarizing voltage steps(from +64 to −196 mV; in −20 mV increments; hv = −36 mV) and in the absence of Mg-ATP using eitherK+ (30 mM) or Ba2+ (100 mM) as charge carriers. Notice the larger IBa compared to IK,in. (B) Current(left panel) and corresponding I-V relationships (right panel) recorded in the same external conditionsas in (A), but this time using only Mg2+ ions in the patch solution (1 mM added as MgCl2) and noadded ATP. In these conditions, the predominant current activated by hyperpolarization is IK,in, not IBa.
Furthermore, and as a control for the “ion transport functionality” of the patched GCPs, we usedconditions that allow measurements of not just IBa, but also to record IK,in. The experiments werestarted in conditions allowing to probe for IK,in with KCl (30 mM) in the bath, which was then followedby replacing the KCl with a solution containing BaCl2 (100 mM). This was done first in the absence ofboth intracellular Mg2+ and ATP (Figure 6A), and then repeated on another batch of guard cells withinternal medium containing 1 mM MgCl2 but no ATP (Figure 6B). Firstly, in zero Mg-ATP, the onlycurrent that activated in response to hyperpolarization was IBa (Figure 6A). IK,in could not be activated.Secondly, when only Mg2+ was included in the pipet solution (no ATP added), IK,in could then beactivated in 30 mM K+. However, when switching the bath from K+- to Ba2+-containing media, IBa
currents vanished, indicating that including only Mg2+ in the patch pipet can cause the blockage of IBa
at voltages where it is normally activated in 0 Mg-ATP.
3.7. Can cAMP Activate HACCs in V. faba Guard Cells Despite the Presence of Intracellular Mg2+?
We know from our own previous work [15] that cAMP activated a Gd3+-sensitive HACC in guardcells while recording in Mg2+- and ATP-free media (Figure 7A). However, since we had shown that thisconductance discriminates poorly between divalent and monovalent cations, a hallmark characteristicof all animal and plant cyclic nucleotide gated channels (CNGCs) [13,45], we sought to check if the

Int. J. Mol. Sci. 2020, 21, 3771 11 of 19
Gd3+-sensitive IBa is also gated by cAMP in conditions where intracellular Mg2+ is present and IBa isalready blocked (Figure 7B).
Int. J. Mol. Sci. 2020, 21, x 11 of 19
characteristic of all animal and plant cyclic nucleotide gated channels (CNGCs) [13,45], we sought to check if the Gd3+-sensitive IBa is also gated by cAMP in conditions where intracellular Mg2+ is present and IBa is already blocked (Figure 7B).
Figure 7. Dibutyryl 3',5'-cyclic adenosine monophosphate (cAMP) potentiates a Gd3+-sensitive current in guard cells either in the presence or absence of intracellular Mg2+. Experiments were conducted in the whole cell configuration, where the GCPs were held at -52 mV and the I-V plots were generated using a hyperpolarizing ramp protocol. See Methods for bath and intracellular media. (A) Typical example from a V. faba GCP patched with no Mg-ATP in the intracellular media, showing superimposed I-V ramps from +50 to −156 mV (70 mV.s−1) in the absence (- db2cAMP) or presence (+db2cAMP) of 1 mM dibutyryl cAMP. Note that adding 0.05 mM GdCl3 while keeping db2cAMP in the bath blocks this conductance. (B) A representative example from an A. thaliana GCP patched with 1mM MgCl2 (no added ATP) in the intracellular media, showing superimposed I-V ramps from ~50 to −192 mV (70 mV.s−1) in the absence (−db2cAMP) or presence (+db2cAMP) of 1 mM dibutyryl cAMP. Note that adding 0.05 mM GdCl3 while keeping db2cAMP in the bath also blocks this conductance.
In the absence of db2cAMP (the lipophilic permeable analog of cAMP), and as expected only a small background IBa current is seen (≤ 10 pA around −190 mV). After perfusing with db2cAMP (1 mM), a substantial increase in IBa amplitude between voltages from around −30 to −190 mV was observed (>60 pA around −190 mV). Keeping db2cAMP in the bath and adding Gd3+ (50 µM) resulted in a total block of the current (≤ 4 pA around −190 mV). This may indicate that GCPs harbor CNGCs that can be activated by cAMP despite the blocking effect by Mg2+.
3.8. Identification of Candidate Mg2+-Dependent Cation Channels in Arabidopsis thaliana
Given the data from the physiological experiments, we undertook a search for candidate Mg2+-dependent cation channels in plants (Figure 8).
A
B
-150 -100 -5050
-400
-200
V (mV)
I (pA)
+ db2cAMP + Gd3+
- db2cAMP
+ db2cAMP
0 MgCl2 (no ATP)
V (mV)
-200 -150 -100 -5050
I (pA)-50
+ db2cAMP + Gd3+
- db2cAMP
+ db2cAMP
1mM MgCl2 (no ATP)
Figure 7. Dibutyryl 3′,5′-cyclic adenosine monophosphate (cAMP) potentiates a Gd3+-sensitive currentin guard cells either in the presence or absence of intracellular Mg2+. Experiments were conducted inthe whole cell configuration, where the GCPs were held at -52 mV and the I-V plots were generatedusing a hyperpolarizing ramp protocol. See Methods for bath and intracellular media. (A) Typicalexample from a V. faba GCP patched with no Mg-ATP in the intracellular media, showing superimposedI-V ramps from +50 to −156 mV (70 mV.s−1) in the absence (−db2cAMP) or presence (+db2cAMP) of 1mM dibutyryl cAMP. Note that adding 0.05 mM GdCl3 while keeping db2cAMP in the bath blocksthis conductance. (B) A representative example from an A. thaliana GCP patched with 1mM MgCl2(no added ATP) in the intracellular media, showing superimposed I-V ramps from ~50 to −192 mV(70 mV.s−1) in the absence (−db2cAMP) or presence (+db2cAMP) of 1 mM dibutyryl cAMP. Note thatadding 0.05 mM GdCl3 while keeping db2cAMP in the bath also blocks this conductance.
In the absence of db2cAMP (the lipophilic permeable analog of cAMP), and as expected onlya small background IBa current is seen (≤10 pA around −190 mV). After perfusing with db2cAMP(1 mM), a substantial increase in IBa amplitude between voltages from around −30 to −190 mV wasobserved (>60 pA around −190 mV). Keeping db2cAMP in the bath and adding Gd3+ (50 µM) resultedin a total block of the current (≤4 pA around −190 mV). This may indicate that GCPs harbor CNGCsthat can be activated by cAMP despite the blocking effect by Mg2+.
3.8. Identification of Candidate Mg2+-Dependent Cation Channels in Arabidopsis thaliana
Given the data from the physiological experiments, we undertook a search for candidateMg2+-dependent cation channels in plants (Figure 8).

Int. J. Mol. Sci. 2020, 21, 3771 12 of 19Int. J. Mol. Sci. 2020, 21, x 12 of 19
Figure 8. Structural rationale for the channel-blocking effect of Mg2+. (A) Sequence alignment of the AtCNGC protein sequence surrounding the putative Mg2+ binding site. The diacidic motif present in most AtCNGCs is highlighted by a red arrow and black line, whereas the location corresponding to the Mg2+ binding acidic motif in the Kir2.2 pore domain is indicated by the left arrow (grey filling). The residues of the pore and linker regions are underlined in green and yellow, respectively. (B) Structural model of AtCNGC8 based on human HCN1 (PDB accession number 5u6o; for the sequence: see Figure S2A). Top panel: side view, with the transmembrane region colored in green, the linker region in yellow, and the cytoplasmic cAMP binding domain in grey. The diacidic motifs (residues E437 and E438 in AtCNGC8) are highlighted as red sphere models. Bottom panel: view from the cytoplasm into the channel (90˚ rotation with respect to top panel). (C) Electrostatic surface representation (color-ramped from negatively charged in red to positively charged in blue) of homology models of AtCNGCs containing the diacidic motif (AtCNGC8 and 6) and of AtCNGC2 (model) and human HCN1 (PDB 5u6o) lacking this motif.
Crystallographic and functional analyses of a strong inward-rectifying K+ channel (Kir2.2) from chicken [46] showed that the rectification characteristic can be explained by Mg2+ binding to negatively charged regions in the pore (formed by D173), with possible additional contributions from acidic residues within the cytoplasmic regulatory domains (D256 and E300/E225) (Figure S2). To explore the possibility that a similar mechanism allows Mg2+ inward rectification for a subset of candidate plant channels that display the electrophysiological properties described here (i.e., activation by hyperpolarization and cAMP, absence of cation selectivity, and inhibition by lanthanides), we built homology models of the pore-forming residues for AtCNGCs [47,48]. The models were built using structures of the human hyperpolarization-activated channels HCN1, based on a ~22% sequence identity. The obtained models clearly showed that AtCNGCs do not have a Mg2+
A
B C
AtCNGC8
AtCNGC6
AtCNGC2
HCN1
Figure 8. Structural rationale for the channel-blocking effect of Mg2+. (A) Sequence alignment of theAtCNGC protein sequence surrounding the putative Mg2+ binding site. The diacidic motif present inmost AtCNGCs is highlighted by a red arrow and black line, whereas the location corresponding to theMg2+ binding acidic motif in the Kir2.2 pore domain is indicated by the left arrow (grey filling). Theresidues of the pore and linker regions are underlined in green and yellow, respectively. (B) Structuralmodel of AtCNGC8 based on human HCN1 (PDB accession number 5u6o; for the sequence: see FigureS2A). Top panel: side view, with the transmembrane region colored in green, the linker region inyellow, and the cytoplasmic cAMP binding domain in grey. The diacidic motifs (residues E437 andE438 in AtCNGC8) are highlighted as red sphere models. Bottom panel: view from the cytoplasminto the channel (90 ◦ rotation with respect to top panel). (C) Electrostatic surface representation(color-ramped from negatively charged in red to positively charged in blue) of homology models ofAtCNGCs containing the diacidic motif (AtCNGC8 and 6) and of AtCNGC2 (model) and human HCN1(PDB 5u6o) lacking this motif.
Crystallographic and functional analyses of a strong inward-rectifying K+ channel (Kir2.2) fromchicken [46] showed that the rectification characteristic can be explained by Mg2+ binding to negativelycharged regions in the pore (formed by D173), with possible additional contributions from acidicresidues within the cytoplasmic regulatory domains (D256 and E300/E225) (Figure S2). To explorethe possibility that a similar mechanism allows Mg2+ inward rectification for a subset of candidateplant channels that display the electrophysiological properties described here (i.e., activation byhyperpolarization and cAMP, absence of cation selectivity, and inhibition by lanthanides), we builthomology models of the pore-forming residues for AtCNGCs [47,48]. The models were built usingstructures of the human hyperpolarization-activated channels HCN1, based on a ~22% sequence

Int. J. Mol. Sci. 2020, 21, 3771 13 of 19
identity. The obtained models clearly showed that AtCNGCs do not have a Mg2+ binding sitecorresponding to the location of Kir2.2 D173 inside the pore region (Figure 8A, left arrow, and Figure S2).However, a tandem glutamic acid motif that could form a possible diacidic Mg2+ binding site is foundin most AtCNGCs located a little downstream of the pore, in the so-called C-linker disc (Figure 8). Akinto Kir2.2 D173 and E300/E225, this diacidic AtGNCG motif forms a positively charged opening, withdistances of 7.3–8.3Å between neighbouring charges and a diameter of ~11Å (for Kir2.2 D173, thesevalues are 7.3–7.4Å and 10.4–11.8Å, respectively (PDB 3jyc)). As in Kir2.2, these distances betweencarboxyl groups are too large for direct ion coordination, suggesting that Mg2+ is bound throughbridging water molecules [46]. Interestingly, this diacidic motif is not present in AtGNCG2, which hasbeen shown to be an atypical family member with respect to ion selectivity [49]. The diacidic motif isalso absent in HCN1, for which Mg2+ inward rectification has not been documented (Figure 8C).
4. Discussion
In order to record ion currents (for instance IK,in or IK,out) from plant cells in the “whole-cell”patch configuration, for reasons highlighted in the introduction (also ass Methods section in [50]), it isstandard to include ATP and Mg2+ in either the form of [Mg-ATP] or [MgCl2 + K2-ATP]. In contrast, thecomposition of the internal solution used to characterize the hyperpolarization-activated Ca2+ current(HACC or IBa) is more variable, especially with regards to ATP and Mg2+. For example, one can noticethat ATP and Mg2+ are either both included [9,11,17,51,52] or completely omitted from the internalsolution [10,16,53–55]. Here, we summarize the role and consequences of Mg2+ inclusion, either in theform of Mg-ATP or MgCl2. Incidentally, there are no data on cytosolic magnesium concentrations orthe distribution of magnesium within the different cellular compartments of guard cells. However, wethink it is reasonable to assume that intracellular free [Mg2+] is between 400 and 600 µM, which thetotal magnesium, including the complexed form, may be as high as 10 mM.
An overview of some of the pharmacological properties related to the HACC (IBa) and IK,in arepresented in Figure 9.
Int. J. Mol. Sci. 2020, 21, x 13 of 19
binding site corresponding to the location of Kir2.2 D173 inside the pore region (Figure 8A, left arrow, and Figure S2). However, a tandem glutamic acid motif that could form a possible diacidic Mg2+ binding site is found in most AtCNGCs located a little downstream of the pore, in the so-called C-linker disc (Figure 8). Akin to Kir2.2 D173 and E300/E225, this diacidic AtGNCG motif forms a positively charged opening, with distances of 7.3–8.3Å between neighbouring charges and a diameter of ~11Å (for Kir2.2 D173, these values are 7.3–7.4Å and 10.4–11.8Å, respectively (PDB 3jyc)). As in Kir2.2, these distances between carboxyl groups are too large for direct ion coordination, suggesting that Mg2+ is bound through bridging water molecules [46]. Interestingly, this diacidic motif is not present in AtGNCG2, which has been shown to be an atypical family member with respect to ion selectivity [49]. The diacidic motif is also absent in HCN1, for which Mg2+ inward rectification has not been documented (Figure 8C).
4. Discussion
In order to record ion currents (for instance IK,in or IK,out) from plant cells in the “whole-cell” patch configuration, for reasons highlighted in the introduction (also ass Methods section in [50]), it is standard to include ATP and Mg2+ in either the form of [Mg-ATP] or [MgCl2 + K2-ATP]. In contrast, the composition of the internal solution used to characterize the hyperpolarization-activated Ca2+ current (HACC or IBa) is more variable, especially with regards to ATP and Mg2+. For example, one can notice that ATP and Mg2+ are either both included [9,11,17,51,52] or completely omitted from the internal solution [10,16,53–55]. Here, we summarize the role and consequences of Mg2+ inclusion, either in the form of Mg-ATP or MgCl2. Incidentally, there are no data on cytosolic magnesium concentrations or the distribution of magnesium within the different cellular compartments of guard cells. However, we think it is reasonable to assume that intracellular free [Mg2+] is between 400 and 600 µM, which the total magnesium, including the complexed form, may be as high as 10 mM.
An overview of some of the pharmacological properties related to the HACC (IBa) and IK,in are presented in Figure 9.
Figure 9. Overview of the properties of the hyperpolarization-activated calcium channels (HACCs).
We report experiments designed to assess the effect of Mg2+ on IBa current activated by hyperpolarization in GCPs, being aware that other cell types, e.g., root cells, might indeed show different responses. We show that omitting Mg-ATP unmasked the presence of an instantaneously activating, inwardly rectifying conductance. This “newly unveiled” conductance still retains most of the biophysical and pharmacological characteristics that are hallmarks of the classical ICa-type [9,10] and CNGCs, which we referred to here as either IBa or HACCs [13,14]. Similarly to IK,in, this conductance is activated by hyperpolarizing-going voltages, and shows some voltage-dependent rectification (but not as strongly as compared to IK,in, for example; see Figure 4A). Additionally, typical of HACCs, we found that besides Ba2+, other divalent cations such as Ca2+, Sr2+, and Mn2+ also permeate this channel, but at a much slower rate than Ba2+ does, hence the smaller currents resolved even though the same amounts of divalent cations in the bath were used. More importantly, this
Extracellular
Activation by hyperpolarization: Gd3+ Mn2+ Ba2+ Cs+ TEA+ Ca2+ Mg2+ ATP cAMP/
cGMP
IBa
Yes Yes Yes
(Véry & Davies, 2000)
No No No Yes
(Hamilton et al., 2000)
Yes
(decreases with high Mg2+)
No Yes
(cAMP potentiates; Lemtiri &
Berkowitz, 2003)
IK,in
Yes No No Yes Yes Yes
(Blatt, 1992)
Yes
(Schroeder & Hagiwara,
1989)
Yes
(increases with high Mg2+)
No
(can restore IKin run down;
Hoshi, 1995)
Yes
(cGMP inhibits; Hoshi, 1995)
Intracellular
Figure 9. Overview of the properties of the hyperpolarization-activated calcium channels (HACCs).
We report experiments designed to assess the effect of Mg2+ on IBa current activated byhyperpolarization in GCPs, being aware that other cell types, e.g., root cells, might indeed showdifferent responses. We show that omitting Mg-ATP unmasked the presence of an instantaneouslyactivating, inwardly rectifying conductance. This “newly unveiled” conductance still retains most of thebiophysical and pharmacological characteristics that are hallmarks of the classical ICa-type [9,10] andCNGCs, which we referred to here as either IBa or HACCs [13,14]. Similarly to IK,in, this conductanceis activated by hyperpolarizing-going voltages, and shows some voltage-dependent rectification (butnot as strongly as compared to IK,in, for example; see Figure 4A). Additionally, typical of HACCs,we found that besides Ba2+, other divalent cations such as Ca2+, Sr2+, and Mn2+ also permeate this

Int. J. Mol. Sci. 2020, 21, 3771 14 of 19
channel, but at a much slower rate than Ba2+ does, hence the smaller currents resolved even thoughthe same amounts of divalent cations in the bath were used. More importantly, this HACC is activatedby cAMP, as well as permeable to monovalent cations (see Figure S1), which are definite attributes ofCNGCs. HACCs, including CNGCs, are specifically blocked by low concentrations of extracellularGd3+ that are far less effective in blocking IK,in (see Figure 4A). Equally important, we found that theunmasked IBa is also enhanced by ≈1.3- to 1.5-fold in response to 5 min treatment with ABA and by upto 2-fold after 10 min. Similarly, it was shown that ABA increases a PM ICa-type whole-cell currentin Arabidopsis guard cells by ≈2- to 3-fold within 5 min of treatment [9]. Note that the ABA effectreported here was obtained not only in Mg2+-free internal solution, but even more importantly inATP-free internal solution, thus indicating that ATP is not as crucial for this channel as was suggestedby an earlier report [56]. This is in agreement with many other reports showing that ABA can indeedincrease cytosolic Ca2+ levels through activation of PM calcium channels via hyperpolarization inArabidopsis thaliana guard cells [10,57,58]. Furthermore, ATP was neither required for cGMP- norcAMP-activated [49,50] Ca2+-permeable cation channels in many different plant cell types (mesophyll,guard cells, or pollen tubes) that also show many of the HACC characteristics. This observation isin contrast to the above-mentioned report, where ATP and subsequent protein (de)phosphorylationwas described as a prerequisite for an ABA effect on calcium channels (56). This indicates that theremay be more than one subtype of calcium channel co-existing in the PM or that additional modesof regulation of these Ca2+-permeable channels are in operation, which might require ATP- andprotein-kinase-dependent signalling [56,59].
One discrepancy that stands out in our report is that Mg2+ was shown to permeate HACCs inguard cells [10] and in root hairs [51], whereas in our experiments external Mg2+ did not appear topermeate this channel (Figure 3A). This is even more intriguing when considering that cGMP wasrecently shown to activate an inward rectifying current (which was also lanthanide-sensitive), withMg2+ as a charge carrier [16]. Hence, this is another hint that we may be dealing with more than onesubtype of calcium channel. In animal cells, Mn2+ was described as a blocker of calcium channelsif Ca2+ is present in the bath, while in the absence of Ca2+ the Mn2+ permeates the channel [60,61].These data might infer that HACCs, despite sharing many similarities in terms of their biophysical andpharmacological characteristics, might slightly differ from one cell type to another depending on thetissue type or plant species.
Therefore, the first key finding is that omitting Mg-ATP from the intracellular medium unmasksa larger Gd3+-sensitive, non-selective cation conductance that is also regulated by cAMP and ABA.The mechanism consists of shifting the I-V characteristic to the right, where less negative voltages canmobilize cations (including Ca2+) through the channel. The second key finding is the demonstrationthat Mg2+ alone can block this conductance (Figure 6).
The current activated by voltages lower than −200 mV is small but significant (Figure 1A). Atthe present time, we have no evidence to support that this instantaneous, rectifying, Mg2+-resistantBa2+ current would be carried by a different population type of HACCs (the channel type that wasunmasked when Mg2+ was omitted). If anything, this current could still be carried by the same typeof channel, since addition of 20 µM Gd3+ to the bath was still able to swiftly and efficiently blockthis current.
Our data describe that in guard cells, Mg2+ blocks IBa by shifting the I-V relationship andits activation threshold to more negative voltages (Figure 1C). This effect is reminiscent of theinhibitory effects by Mg2+ on many ion channels, which have been interpreted as “charge screeningeffects” [18]. Indeed, the rectification of the inwardly rectifying K+ (Kir) channels is due to avoltage-dependent blockage by cytosolic Mg2+ (and polyamines), thereby blocking outward K+-efflux.Upon hyperpolarization, Mg2+ is ejected from the pore, which appears to result in a time-dependentopening of the channel [62,63]. Likewise, TRPV6 shows Mg2+-dependent gating that contributes to itsstrong inward rectification [20]; it was suggested that Mg2+ can block the channel by binding to a sitewithin the transmembrane electrical field, where it interacts with other permeant cations [20]. It is also

Int. J. Mol. Sci. 2020, 21, 3771 15 of 19
conceivable that other mechanisms could be operating, such as electrostatic interaction between Mg2+
and some PM lipids, such as phosphatidylinositol 4,5-bisphosphate (or PIP2). It was demonstratedthat increasing the amount of membrane PIP2 results in decreasing the sensitivity of KCNQ channel toinhibition by Mg2+ [64]. In addition, some Ca2+ channels were found to require PIP2 for their normalfunction [65]. This begs the question of whether guard cell PM Ca2+ channels are also PIP2-sensitive.Our results also raise the question of whether Mg2+ could be equally important for IK,in gating. Indeed,unlike earlier reports, we found that activation of IK,in was dependent on Mg2+ being present insidethe patch pipet (Figure 6).
Furthermore, the activation of the above HACCs by hyperpolarization and cAMP, the absenceof cation selectivity, and inhibition by lanthanides is consistent with the hypothesis that channelsresponsible for the observed effects are CNGCs. This is consistent with our structural modelling,which revealed the presence of diacidic motifs in the pore-forming helix of a subset of AtCNGCs.Acidic residues pointing towards the inner side of the pore, have previously been shown to conferMg2+ dependence to inward rectifying K+ channels in animals [47] (Figure S2B). Given the positionof the diacidic motif in the cytoplasmic side of the pore, it is conceivable that Mg2+ binding can beaffected by changes in pore opening, for example introduced by cAMP binding to the cytoplasmicregion of AtCNGCs. Such crosstalk would provide a mechanistic explanation for our observation thatcAMP overrides the channel blockage produced by 1mM MgCl2 (no added ATP) (Figure S2C), therebypreventing uncontrolled Ca2+ leakage.
5. Conclusions
Here, we firstly propose that in Vicia faba guard cells, Mg2+ can limit or prevent continuousCa2+-leakage, possibly through all HACCs (including CNGCs) at resting membrane potentials, therebybeing part of the (intra)cellular calcium signaling processes [66]. Furthermore, CNGCs have also beenrecognized as having a critical role in Ca2+-dependent plant defense signalling and responses [67,68].Secondly, we propose that the activation of these channels requires mechanism(s) by which Mg2+
binding is altered. In CNGCs, such a mechanism is conceivably enabled by cAMP binding, therebyassigning Mg2+ an important role in calcium homeostasis and calcium-dependent downstreamprocesses. Finally, the effect reported here suggests that Mg2+ has a role in cellular Ca2+ homeostasis,similarly to in animal cells.
Supplementary Materials: Supplementary materials can be found at http://www.mdpi.com/1422-0067/21/11/3771/s1: Supplementary Figure S1: In the absence of MgATP, guard cell HACCs are permeable to monovalentcations, such as K+, Na+, and Cs+, but not TEA+. All experiments were conducted in the whole cell configuration,where V. faba GCPs were held at −56 mV. (A) Superimposed I-V plots in the presence of 100 mM BaCl2 (�), 100mM KCl (N), or 100 KCl + 0.05 mM GdCl3. (B) Superimposed I-V plots in the presence of 100 mM BaCl2 (�), 100mM NaCl (l), or 100 NaCl + 0.05 mM GdCl3. (C) Superimposed I-V plots in the presence of 100 mM BaCl2 (�) or100 mM CsCl (�), or 100 CsCl + 0.05 mM GdCl3 (−). (D) Superimposed I-V plots in the presence of 100 mM BaCl2(�) or 100 TEACl (H). Supplementary Figure S2: (A) Alignment of the Arabidopsis thaliana CNGC8 pore region andthe Kir2.2 (PDB accession number 5u6o_341_471). (B) Mechanism of inward rectification by magnesium ions.Tao et al. (Science 2009, 326, 1668-1674) have shown that inward rectification through Mg2+ can be explained bythe ion binding to negatively charged regions in the pore (formed by D173) and in the cytoplasmic regulatorydomains (D256 and E300/E225). The crystal structure of the inward rectifying potassium channel Kir2.2 (Tao,2009; PDB entry3JYC) is shown in ribbon presentation. The four subunits are color-coded. Potassium ions in thechannel are shown as magenta spheres. Negatively charged residues that bind the Mg2+ mimic Sr2+ in the crystalstructure are shown in pink on their molecular surfaces. (C) Sequences of pore-forming transmembrane helix(TM) from AtCNGCs. The TM is marked with a dashed line. Models were built using Swiss-Model (Waterhouseet al. (Nucleic Acids Res. 2018, 46, W296-W303)).
Author Contributions: F.L.-C. and C.G. conceived the study. F.L.-C. performed the experiments and analyzedthe data. C.G. and S.T.A. performed the structural analyses. All authors have read and agreed to the publishedversion of the manuscript.
Funding: This research has been supported by the King Abdullah University of Science and Technology (KAUST).We are indebted to Enid MacRobbie (Department of Plant Science, University of Cambridge, UK) for allowing usto use some of the data gathered by F.L.-C. whilst in her laboratory (research was supported by BBSRC GrantP05730 to E.M.). We also thank Mark Tester for his invaluable comments.

Int. J. Mol. Sci. 2020, 21, 3771 16 of 19
Acknowledgments: We also thank Mark Tester for his invaluable comments.
Conflicts of Interest: The authors declare no conflict of interest.
References
1. McAinsh, M.R.; Pittman, J.K. Shaping the calcium signature. New Phytol. 2009, 181, 275–294. [CrossRef][PubMed]
2. Véry, A.A.; Sentenac, H. Cation channels in the Arabidopsis plasma membrane. Trends Plant Sci. 2002, 7,168–175. [CrossRef]
3. White, P.J.; Broadley, M.R. Calcium in plants. Ann. Bot. 2003, 92, 487–511. [CrossRef] [PubMed]4. Demidchik, V.; Davenport, R.J.; Tester, M. Nonselective cation channels in plants. Annu. Rev. Plant Biol. 2002,
53, 67–107. [CrossRef]5. Thion, L.; Mazars, C.; Nacry, P.; Bouchez, D.; Moreau, M.; Ranjeva, R.; Thuleau, P. Plasma membrane
depolarization-activated calcium channels, stimulated by microtubule-depolymerizing drugs in wild-typeArabidopsis thaliana protoplasts, display constitutively large activities and a longer half-life in ton 2 mutantcells affected in the organization of cortical microtubules. Plant J. 1998, 13, 603–610.
6. Miedema, H.; Bothwell, J.H.; Brownlee, C.; Davies, J.M. Calcium uptake by plant cells - channels and pumpsacting in concert. Trends Plant Sci. 2001, 6, 514–519. [CrossRef]
7. MacRobbie, E.A.C. Calcium and ABA-induced stomatal closure. Philos. Trans. Roy. Soc. B. 1992, 338, 5–18.8. MacRobbie, E.A.C. ABA activates multiple Ca2+ fluxes in stomatal guard cells, triggering vacuolar K+(Rb+)
release. Proc. Natl. Acad. Sci. USA 2000, 97, 12361–12368. [CrossRef]9. Hamilton, D.W.A.; Hills, A.; Köhler, B.; Blatt, M.R. Ca2+ channels at the plasma membrane of stomatal guard
cells are activated by hyperpolarization and abscisic acid. Proc. Natl. Acad. Sci. USA 2000, 97, 4967–4972.[CrossRef]
10. Pei, Z.M.; Murata, Y.; Benning, G.; Thomine, S.; Klusener, B.; Allen, G.J.; Grill, E.; Schroeder, J.I. Calciumchannels activated by hydrogen peroxide mediate abscisic acid signalling in guard cells. Nature 2000, 406,731–734. [CrossRef]
11. Hamilton, D.W.A.; Hills, A.; Blatt, M.R. Extracellular Ba2+ and voltage interact to gate Ca2+ channels at theplasma membrane of stomatal guard cells. FEBS Lett. 2001, 491, 99–103. [CrossRef]
12. Leng, Q.; Mercier, R.W.; Yao, W.; Berkowitz, G.A. Cloning and first functional characterization of a plantcyclic nucleotide-gated cation channel. Plant Physiol. 1999, 121, 753–761. [CrossRef] [PubMed]
13. Leng, Q.; Mercier, R.W.; Hua, B.G.; Fromm, H.; Berkowitz, G.A. Electrophysiological analysis of cloned cyclicnucleotide-gated ion channels. Plant Physiol. 2002, 128, 400–410. [CrossRef] [PubMed]
14. Balagué, C.; Lin, B.Q.; Alcon, C.; Flottes, G.; Malmström, S.; Köhler, C.; Neuhaus, G.; Pelletier, G.; Gaymard, F.;Roby, D. HLM1, an essential signaling component in the hypersensitive response, is a member of the cyclicnucleotide-gated channel ion channel family. Plant Cell 2003, 15, 365–379. [CrossRef] [PubMed]
15. Lemtiri-Chlieh, F.; Berkowitz, G.A. Cyclic adenosine monophosphate regulates calcium channels in theplasma membrane of Arabidopsis leaf guard and mesophyll cells. J. Biol. Chem. 2004, 279, 35306–35312.[CrossRef] [PubMed]
16. Wang, Y.F.; Munemasa, S.; Nishimura, N.; Ren, H.M.; Robert, N.; Han, M.; Puzorjova, I.; Kollist, H.; Lee, S.;Mori, I.; et al. Identification of cyclic GMP-activated nonselective Ca2+-permeable cation channels andassociated CNGC5 and CNGC6 genes in Arabidopsis guard cells. Plant Physiol. 2013, 163, 578–590. [CrossRef]
17. Wu, J.Y.; Qu, H.Y.; Jin, C.; Shang, Z.L.; Wu, J.; Xu, G.H.; Gao, Y.B.; Zhang, S.L. cAMP activateshyperpolarization-activated Ca2+-channels in the pollen of Pyrus pyrifolia. Plant Cell Rep. 2011, 30, 1193–1200.[CrossRef] [PubMed]
18. Hille, B. Ion Channels of Excitable Membranes, 3rd ed.; Sinauer Associates, Inc.: Sunderland, MA, USA, 2001.19. Matsuda, H. Open-state substructure of inwardly rectifying potassium channels revealed by magnesium
block in guinea-pig heart-cells. J. Physiol. 1988, 397, 237–25820. [CrossRef]20. Voets, T.; Janssens, A.; Prenen, J.; Droogmans, G.; Nilius, B. Mg2+-dependent gating and strong inward
rectification of the cation channel TRPV6. J. Gen. Physiol. 2003, 121, 245–260. [CrossRef]21. Zito, K.; Scheuss, V. NMDA Receptor Function and Physiological Modulation. In Encyclopedia of Neuroscience;
Squire, L.R., Ed.; Academic Press: Oxford, UK, 2009.

Int. J. Mol. Sci. 2020, 21, 3771 17 of 19
22. Nowak, L.M.; Ascher, P.; Bregestovski, P.; Herbet, A.; Prochiantz, A. Voltage dependence of 1-Glu inducedcurrent is due to gating by Mg ions. Biophys. J. 1984, 45, A388.
23. Schroeder, J.I. Magnesium-independent activation of inward-rectifying K+ channels in Vicia-faba guard-cells.FEBS Lett. 1995, 363, 157–160. [CrossRef]
24. Hedrich, R.; Moran, O.; Conti, F.; Busch, H.; Becker, D.; Gambale, F.; Dreyer, I.; Kuch, A.; Neuwinger, K.;Palme, K. Inward rectifier potassium channels in plants differ from their animal counterparts in response tovoltage and channel modulators. Eur. Biophys. J. 1995, 24, 107–115. [CrossRef]
25. Pei, Z.M.; Ward, J.M.; Schroeder, J.I. Magnesium sensitizes slow vacuolar channels to physiological cytosoliccalcium and inhibits fast vacuolar channels in fava bean guard cell vacuoles. Plant Physiol. 1999, 121, 977–986.[CrossRef]
26. Brüggemann, L.I.; Pottosin, I.I.; Schönknecht, G. Cytoplasmic magnesium regulates the fast activatingvacuolar cation channel. J. Exp. Bot. 1999, 50, 1547–1552. [CrossRef]
27. Carpaneto, A.; Cantu, A.M.; Gambale, F. Effects of cytoplasmic Mg2+ on slowly activating channels inisolated vacuoles of Beta vulgaris. Planta 2001, 213, 457–468. [CrossRef] [PubMed]
28. Pottosin, I.; Martinez-Estevez, M.; Dobrovinskaya, O.; Muniz, J.; Schönknecht, G. Mechanism of luminalCa2+ and Mg2+ action on the vacuolar slowly activating channels. Planta 2004, 219, 1057–1070. [CrossRef][PubMed]
29. Wolf, T.; Guinot, D.R.; Hedrich, R.; Dietrich, P.; Marten, I. Nucleotides and Mg2+ ions differentially regulateK+ channels and non-selective cation channels present in cells forming the stomatal complex. Plant CellPhysiol. 2005, 46, 1682–1689. [CrossRef]
30. Tyerman, S.D.; Whitehead, L.F.; Day, D.A. A channel-like transporter for NH4+ on the symbiotic interface of
N2-fixing plants. Nature 1995, 378, 629–632. [CrossRef]31. Obermeyer, G.; Tyerman, S.D. NH4
+ currents across the peribacteroid membrane of soybean. Macroscopicand microscopic properties, inhibition by Mg2+, and temperature dependence indicate a subpicosiemenschannel finely regulated by divalent cations. Plant Physiol. 2005, 139, 1015–1029. [CrossRef]
32. Babenko, A.P.; Aguilar-Bryan, L.; Bryan, J. A view of SUR/KIR6.X, KATP channels. Annu. Rev. Physiol. 1998,60, 667–687. [CrossRef]
33. Campbell, J.D.; Sansom, M.S.P.; Ashcroft, F.M. Potassium channel regulation - Structural insights into thefunction of the nucleotide-binding domains of the human sulphonylurea receptor. EMBO Rep. 2003, 4,1038–1042. [CrossRef] [PubMed]
34. Assmann, S.M.; Simoncini, L.; Schroeder, J.I. Blue-light activates electrogenic ion pumping in guard-cellprotoplasts of Vicia faba. Nature 1985, 318, 285–287. [CrossRef]
35. Schroeder, J.I. K+ Transport-properties of K+ channels in the plasma-membrane of Vicia faba guard-cells. J.Gen. Physiol. 1988, 92, 667–683. [CrossRef]
36. Blatt, M.R. Electrical characteristics of stomatal guard-cells—The ionic basis of the membrane-potential andthe consequence of KCl leakage from microelectrodes. Planta 1987, 170, 272–287. [CrossRef] [PubMed]
37. Lemtiri-Chlieh, F. Effects of internal K+ and ABA on the voltage- and time-dependence of the outwardK+-rectifier in Vicia guard cells. J. Memb. Biol. 1996, 153, 105–116. [CrossRef] [PubMed]
38. Lemtiri-Chlieh, F.; Macrobbie, E.A.C. Role of calcium in the modulation of Vicia guard-cell potassiumchannels by abscisic acid: A patch-clamp study. J. Memb. Biol. 1994, 137, 99–107. [CrossRef]
39. Neher, E. Correction for liquid junction potentials in patch clamp experiments. Methods Enzymol. 1992, 207,123–131.
40. Shaff, J.E.; Schultz, B.A.; Craft, E.J.; Clark, R.T.; Kochian, L.V. GEOCHEM-EZ: A chemical speciation programwith greater power and flexibility. Plant Soil 2010, 330, 207–214. [CrossRef]
41. Waterhouse, A.; Bertoni, M.; Bienert, S.; Studer, G.; Tauriello, G.; Gumienny, R.; Heer, F.T.; de Beer, T.A.P.;Rempfer, C.; Bordoli, L.; et al. SWISS-Model: Homology modelling of protein structures and complexes.Nucleic Acids Res. 2018, 46, W296–W303. [CrossRef]
42. Buchan, D.W.; Minneci, F.; Nugent, T.C.; Bryson, K.; Jones, D.T. Scalable web services for the PSIPREDProtein Analysis Workbench. Nucleic Acids Res. 2013, 41, W349–W357. [CrossRef] [PubMed]
43. Edgar, R.C. MUSCLE: A multiple sequence alignment method with reduced time and space complexity.BMC Bioinform. 2004, 5, 113. [CrossRef]
44. Blatt, M.R. K+ channels of stomatal guard-cells—Characteristics of the inward rectifier and its control by pH.J. Gen. Physiol. 1992, 99, 615–644. [CrossRef] [PubMed]

Int. J. Mol. Sci. 2020, 21, 3771 18 of 19
45. Zagotta, W.N.; Siegelbaum, S.A. Structure and function of cyclic nucleotide-gated channels. Ann. Rev.Neurosci. 1996, 19, 235–263. [CrossRef]
46. Tao, X.; Avalos, J.L.; Chen, J.; MacKinnon, R. Crystal structure of the eukaryotic strong inward-rectifier K+
channel Kir2.2 at 3.1 Å resolution. Science 2009, 326, 1668–1674. [CrossRef] [PubMed]47. Zelman, A.K.; Dawe, A.; Gehring, C.; Berkowitz, G.A. Evolutionary and structural perspectives of plant
cyclic nucleotide-gated cation channels. Front. Plant Sci. 2012, 3, 95. [CrossRef] [PubMed]48. Clayton, G.M.; Altieri, S.; Heginbotham, L.; Unger, V.M.; Morais-Cabral, J.H. Structure of the transmembrane
regions of a bacterial cyclic nucleotide-regulated channel. Proc. Natl. Acad. Sci. USA 2008, 105, 1511–1515.[CrossRef] [PubMed]
49. Hua, B.G.; Mercier, R.W.; Leng, Q.; Berkowitz, G.A. Plants do it differently. A new basis for potassium/sodiumselectivity in the pore of an ion channel. Plant Physiol. 2003, 132, 1353–1361. [CrossRef]
50. Bush, D.S.; Hedrich, R.; Schroeder, J.I.; Jones, R.L. Channel-mediated K+ flux in barley aleurone protoplasts.Planta 1988, 176, 368–377. [CrossRef]
51. Véry, A.A.; Davies, J.M. Hyperpolarization-activated calcium channels at the tip of Arabidopsis root hairs.Proc. Natl. Acad. Sci. USA 2000, 97, 9801–9806. [CrossRef] [PubMed]
52. Miedema, H.; Demidchik, V.; Véry, A.A.; Bothwell, J.H.F.; Brownlee, C.; Davies, J.M. Two voltage-dependentcalcium channels co-exist in the apical plasma membrane of Arabidopsis thaliana root hairs. New Phytol. 2008,179, 378–385. [CrossRef]
53. Murata, Y.; Pei, Z.M.; Mori, I.C.; Schroeder, J. Abscisic acid activation of plasma membrane Ca2+ channelsin guard cells requires cytosolic NAD(P)H and is differentially disrupted upstream and downstream ofreactive oxygen species production in abi1-1 and abi2-1 protein phosphatase 2C mutants. Plant Cell 2001, 13,2513–2523. [CrossRef] [PubMed]
54. Demidchik, V.; Bowen, H.C.; Maathuis, F.J.M.; Shabala, S.N.; Tester, M.A.; White, P.J.; Davies, J.M. Arabidopsisthaliana root non-selective cation channels mediate calcium uptake and are involved in growth. Plant J. 2002,32, 799–808. [CrossRef] [PubMed]
55. Lemtiri-Chlieh, F.; MacRobbie, E.A.C.; Webb, A.A.R.; Manison, N.F.; Brownlee, C.; Skepper, J.N.; Chen, J.;Prestwich, G.D.; Brearley, C.A. Inositol hexakisphosphate mobilizes an endomembrane store of calcium inguard cells. Proc. Natl. Acad. Sci. USA 2003, 100, 10091–10095. [CrossRef] [PubMed]
56. Köhler, B.; Blatt, M.R. Protein phosphorylation activates the guard cell Ca2+ channel and is a prerequisite forgating by abscisic acid. Plant J. 2002, 32, 185–194. [CrossRef] [PubMed]
57. Munemasa, S.; Oda, K.; Watanabe-Sugimoto, M.; Nakamura, Y.; Shimoishi, Y.; Murata, Y. Thecoronatine-insensitive 1 mutation reveals the hormonal signaling interaction between abscisic acid andmethyl jasmonate in arabidopsis guard cells. Specific impairment of ion channel activation and secondmessenger production. Plant Physiol. 2007, 143, 1398–1407. [CrossRef]
58. Vahisalu, T.; Kollist, H.; Wang, Y.F.; Nishimura, N.; Chan, W.Y.; Valerio, G.; Lamminmäki, A.; Brosche, M.;Moldau, H.; Desikan, R.; et al. SLAC1 is required for plant guard cell S-type anion channel function instomatal signalling. Nature 2008, 452, 487–491. [CrossRef]
59. Mori, I.C.; Murata, Y.; Yang, Y.; Munemasa, S.; Wang, Y.F.; Andreoli, S.; Tiriac, H.; Alonso, J.M.; Harper, J.F.;Ecker, J.R.; et al. CDPKs CPK6 and CPK3 function in ABA regulation of guard cell S-type anion- andCa2+-permeable channels and stomatal closure. PLoS Biol. 2006, 4, e327. [CrossRef]
60. Coraboeuf, E. Voltage clamp studies of the slow inward current. In The Slow Inward Current and CardiacArrhythmias, 1st ed.; Martinus Nijhoff: The Hague, The Netherlands, 1980; pp. 25–95.
61. Bannister, R.A.; Pessah, I.N.; Beam, K.G. The skeletal L-type Ca2+ current is a major contributor toexcitation-coupled Ca2+ entry. J. Gen. Physiol. 2009, 133, 79–91. [CrossRef]
62. Nichols, C.G.; Lopatin, A.N. Inward rectifier potassium channels. Annu. Rev. Physiol. 1997, 59, 171–191.[CrossRef]
63. Guo, D.L.; Ramu, Y.; Klem, A.M.; Lu, Z. Mechanism of rectification in inward-rectifier K+ channels. J Gen.Physiol. 2003, 121, 261–275. [CrossRef] [PubMed]
64. Suh, B.C.; Hille, B. Electrostatic interaction of internal Mg2+ with membrane PIP2 seen with KCNQ K+
channels. J. Gen. Physiol. 2007, 130, 241–256. [CrossRef]65. Gamper, N.; Reznikov, V.; Yamada, Y.; Yang, J.; Shapiro, M.S. Phosphotidylinositol 4,5-bisphosphate signals
underlie receptor-specific Gq/11-mediated modulation of N-type Ca2+ channels. J. Neurosci. 2004, 24,10980–10992. [CrossRef] [PubMed]

Int. J. Mol. Sci. 2020, 21, 3771 19 of 19
66. Stael, S.; Wurzinger, B.; Mair, A.; Mehlmer, N.; Vothknecht, U.C.; Teige, M. Plant organellar calcium signalling:An emerging field. J. Exp. Bot. 2012, 63, 1525–1542. [CrossRef] [PubMed]
67. Ali, R.; Ma, W.; Lemtiri-Chlieh, F.; Tsaltas, D.; Leng, Q.; von Bodman, S.; Berkowitz, G.A. Death don’t haveno mercy and neither does calcium: Arabidopsis CYCLIC NUCLEOTIDE GATED CHANNEL2 and innateimmunity. Plant Cell 2007, 19, 181–195. [CrossRef]
68. Meena, M.K.; Prajapati, R.; Krishna, D.; Divakaran, K.; Pandey, Y.; Reichelt, M.; Mathew, M.K.; Boland, W.;Mithöfer, A.; Vadassery, J. The Ca2+ channel CNGC19 regulates Arabidopsis defense against Spodopteraherbivory. Plant Cell 2019, 31, 1539–1562. [CrossRef]
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open accessarticle distributed under the terms and conditions of the Creative Commons Attribution(CC BY) license (http://creativecommons.org/licenses/by/4.0/).