1997. human cytotrofoblas diferentiation.pdf
TRANSCRIPT

American Journal ofPathology, Vol. 151, No. 6, December 1997Copynight ) American Societyfor Investigative Pathology
Human Cytotrophoblast Differentiation/Invasion IsAbnormal in Pre-Eclampsia
Kee-Hak Lim,* Yan Zhou,t Mary Janatpour,tMichael McMaster,t Kathryn Bass,tSun-Hee Chun,* and Susan J. Fisher*tt§From the Departments of Obstetrics, Gynecology andReproductive Sciences,* Stomatologyt Anatomy,' andPharmaceutical Chemistry,5 University of California SanFrancisco, San Francisco, California
During human placental development, cytotropho-blast stem cells differentiate and invade the uterus.Simultaneously, the cells modulate their expressionof several classes of stage-specific antigens that marktransitions in the differentiation process and play arole in either uterine invasion (integrin cell-extracel-lular matrix receptors and matrix metailoprotein-ase-9) or immune interactions (HLA-G). The preg-nancy disease pre-eclampsia is associated withshallow cytotrophoblast invasion. Previously weshowed, by immunofluorescence localization on pla-cental tissue, that in pre-eclampsia invasive cytotro-phoblasts fail to properly modulate their integrin rep-ertoire. This finding suggests possible abnormalitiesin the differentiation pathway that leads to uterineinvasion. Here we used a culture system that supportsthis differentiation process to compare the differen-tiative and invasive potential of cytotrophoblasts ob-tained from control (n = 8, 22 to 38 weeks) andpre-eclamptic (n = 9, 24 to 38 weeks) placentas. Inculture, the cells from pre-eclamptic placentas failedto properly modulate al integrin and matrix metal-loproteinase-9 expression at the protein and mRNAlevels. Their invasive potential was also greatly re-duced. Likewise, the cells failed to up-regulate HLA-Gprotein and mRNA expression. These results suggestthat defective cytotrophoblast differentiation/inva-sion can have significant consequences to the out-come of human pregnancy (ie, development of pre-eclampsia) and that, by the time delivery becomesnecessary, the defect is not reversed by removing thecells from the maternal environment. (AmJ Pathol1997, 151:1809-1818)
fetal portion of this interface.1' 2 Cytotrophoblast stemcells are attached to the basement membrane that sur-rounds the stromal core of chorionic villi. Many of thesecells differentiate by fusing to form a syncytial layer thatcovers the villus and transports nutrients, wastes, andgases between fetal and maternal blood. At selectedsites, cytotrophoblasts break through the syncytium andform multilayered columns of cells that attach to andinvade the uterus. As a result of interstitial invasion, asubpopulation of cytotrophoblasts (termed extravillous,invasive, or intermediate)3 are normally found throughoutthe entire decidua and the first third of the myometrium.As a result of endovascular invasion, these cells alsoinvade uterine arterioles, replacing the endothelial liningand most of the musculoelastic tissue in the vesselwall.,45 Consequently, uterine arterioles become high-flow, low-resistance vessels that are capable of respond-ing to increases in fetal requirements for maternal bloodas pregnancy progresses.6We hypothesized that fetal cytotrophoblast invasion of
the uterus requires these cells to up-regulate expressionof molecules that function during this process. The mor-phology of the maternal-fetal interface suggests that thecells' ability to acquire an invasive phenotype, whileavoiding maternal immune recognition, is particularly cru-cial. In studying invasiveness, we showed that cytotro-phoblasts normally modulate their expression of severalintegrin cell-extracellular matrix (ECM) receptors7 andMMP-9.6-10 We used an in vitro model of this differentia-tion process to analyze the role these molecules play.9'11The results showed that, during invasion, the cells up-regulated their expression, first of molecules that inhibitinvasion (eg, the integrin a5 fibronectin receptor) andthen of molecules that promote it (eg, the integrin allaminin and collagen (types and IV) receptor; MMP-9).Therefore, correctly balancing these mechanisms is likelyto be critical to normal placental development. In study-ing how these cells evade maternal immune recognition,we showed that the subpopulation of cells that invade theuterus express HLA-G, a very unusual class lb majorhistocompatibility antigen that may protect them fromnatural killer cell lysis.12-14
Formation of the maternal-fetal interface, a consequenceof cytotrophoblast differentiation, is critical to pregnancyoutcome. In humans, two mutually exclusive differentia-tion pathways give rise to morphologically and function-ally distinct trophoblast populations that compose the
Supported by National Institutes of Health grant HD 30367.Accepted for publication September 11, 1997.Address reprint requests to Dr. Susan Fisher, Department of Stomatol-
ogy, HSW 604, 513 Parnassus Avenue, University of California San Fran-cisco, San Francisco, CA 94143-0512.
1809

1810 Lim et alAJP December 1997, Vol. 151, No. 6
We reasoned that pregnancy diseases in which cy-totrophoblast invasion is abnormal at a microscopic levelcould be associated with disruption of invasion at a mo-lecular level, ie, result from an imbalance of the regulatorymechanisms that govern invasion. Previous work sug-gested that pre-eclampsia could be an interesting exam-ple of such a disease. Histological studies of the mater-nal-fetal interface (eg, placental bed biopsies) show that,in pre-eclampsia, interstitial cytotrophoblast invasion isoften shallow and endovascular invasion nearly ab-sent.15-17 Pre-eclampsia is important because it occursfrequently (7% of nulliparous patients) and has seriousconsequences for both the mother (eg, high blood pres-.sure, proteinuria, and edema) and the fetus (intrauterinegrowth retardation). As a result, pre-eclampsia and hy-pertensive diseases of pregnancy are a leading cause ofmaternal death18 and contribute significantly to prema-ture deliveries in the United States.19We began studying cytotrophoblast differentiation in
this syndrome by using immunolocalization techniques todetermine whether differentiating/invading cytotropho-blasts in tissue sections of the maternal-fetal interfacecorrectly switch their integrin repertoire. In normal preg-nancies, cytotrophoblasts that are differentiating alongthe invasive pathway first up-regulate the expression ofintegrin a5/01, then al/f1.7 In pre-eclampsia, invasivecytotrophoblasts do not completely switch their integrinrepertoire. The cells express integrin a5/f31 but not al/11,17 thus displaying an adhesion phenotype our func-tion-perturbation studies suggest could inhibit invasion.11In addition, Redline20 recently showed that, in pregnan-cies complicated by pre-eclampsia, intermediate/inva-sive cytotrophoblasts have an increased proliferative in-dex, as evidenced by increased staining for proliferatingcell nuclear antigen (PCNA) and reduced expression ofhuman placental lactogen, another antigen that is pro-duced as a consequence of the differentiation process.Together, these results suggest that, in pre-eclampsia,cytotrophoblasts start to differentiate along the invasivepathway but cannot complete this process. However,studies performed on tissue sections do not allow us toexamine the differentiative potential of cytotrophoblastsand to determine whether the observed defect is revers-ible.
In experiments described here, we used a culturesystem that promotes cytotrophoblast stem cell differen-tiation along the invasive pathway to compare the differ-entiative potential of cells isolated from placentas of ges-tation-matched control and pre-eclamptic patients. Theresults showed that, in pre-eclampsia, cytotrophoblastintegrin switching in vitro was altered in a pattern thatreplicated the changes we previously described in vivo,ie, in tissue sections of placental bed biopsies. Cytotro-phoblast expression of al integrin protein and mRNAwas all but absent. Their ability to up-regulate MMP-9production, as well as to invade ECM in vitro, was alsoimpaired. Likewise, their expression of HLA-G, a stage-specific antigen that probably plays an important role inmaternal-fetal immune interactions, dramatically de-creased. Together, these data suggest that, in pre-eclampsia, cytotrophoblasts start to differentiate along
the invasive pathway, fail to complete the process, andconsequently are less able to invade. As neither stage-specific antigen expression nor invasiveness could berestored to control levels by removing the cells from thematernal environment, these changes are irreversible bythe time the symptoms necessitate delivery.
Materials and Methods
Placental SourcesCytotrophoblasts were isolated from the placentas of pa-tients with pre-eclampsia diagnosed according to thefollowing criteria, recommended by Chesley21: nullipar-ity, no history of hypertension before pregnancy, increasein diastolic pressure of 15 mm Hg or systolic pressure of30 mm Hg compared with blood pressure obtained be-fore 20 weeks of gestation, proteinuria of .0.5 g/24 hoursor .300 mg/dl (or 1 + on urine dipstick) in a catheterizedspecimen, hyperuricemia of >5.5 mg/dl (or one SDgreater than the normal mean value before term), andreturn to normal blood pressure and resolution of protein-uria by 12 weeks postpartum. The American College ofObstetricians and Gynecologists' definitions for severepre-eclampsia19 and published criteria for the syndromeof hemolysis, elevated liver enzymes, and low platelets(HELLP)22 were used: systolic blood pressure of .160mm Hg and/or diastolic pressure of .110 mm Hg, pro-teinuria of .5 gm in a 24-hour period or 3+ on urinedipstick, presence of cerebral or visual disturbances,epigastric pain, oliguria (<500 ml in 24 hours), pulmonaryedema, evidence of hemolysis on peripheral smear, in-creased bilirubin of .1.2 mg/dl, increased lactic dehy-drogenase of >600 IU/L, platelet counts of <100,000/mm3, and serum glutamic-oxaloacetic transaminase levelof .70 lU/ml. Eclampsia was defined as an onset ofseizure activity in the setting of pre-eclampsia in a patientwithout a history of seizure disorder. Of the nine patientswhose placentas we obtained, one was diagnosed withpre-eclampsia at 38 weeks of gestation, five with severepre-eclampsia at 24, 27, 28, 29, and 30 weeks of gesta-tion, two with HELLP syndrome at 28 and 32 weeks, andone with eclampsia/severe pre-eclampsia at 29 weeks.
Of the eight total control patients, seven patients de-livered at or before 33 weeks. Two delivered at 27 and 32weeks due to cervical incompetence. Three underwentpregnancy terminations at 22 to 24 weeks, and the sixthpatient delivered at 33 weeks because of microinvasivecarcinoma of the cervix diagnosed during pregnancy.The seventh patient underwent elective delivery at 26weeks of gestation for nonoperable conjoined twins.None of the patients had evidence of pre-eclampsia,gestational hypertension, chorioamnionitis, chronic hy-pertension, or a medical history that suggested they wereat an increased risk for developing pre-eclampsia. Noneof the placentas had abnormalities that could be de-tected either grossly or histologically. The eighth patientdelivered at 38 weeks after a normal, uncomplicatedpregnancy.

Abnormal Cytotrophoblast Differentiation/Invasion in Pre-Eclampsia 1811AJP December 1997, Vol. 151, No. 6
Isolation and Culture of CytotrophoblastsCytotrophoblasts were isolated from second-trimesterplacentas according to the methods described previous-ly.8'9 Briefly, chorionic villi were subjected to enzymaticdissociation using trypsin, DNAse 1, and collagenase.The resulting cells were enriched using Percoll gradientcentrifugation. The cytotrophoblast-enriched fractionfrom the Percoll gradient was purified further by removingbone-marrow-derived cells using magnetic beads (Per-Septive Diagnostics, Cambridge, MA) coated with anti-CD45 antibody, which recognizes all bone-marrow-de-rived cells (purified from ascites fluid of the GAP 8.3hybridoma, American Type Culture Collection, Rockville,MD). Cytotrophoblasts from third-trimester placentaswere isolated by similar methods except that the chori-onic villi were subjected to three cycles of enzymaticdigestion using trypsin and DNAse 1. All isolated cytotro-phoblasts were cultured in DME H21 minimal essentialmedium containing 2% Nutridoma (Boehringer Mann-heim, Indianapolis, IN) and 50 ,ug/ml gentamicin. Viabilityafter 72 hours in culture was 90 to 95% as determined bytrypan blue exclusion.
ImmunohistochemistryMouse monoclonal antibody to the human integrin al
subunit (IgG) was obtained from T-Cell Laboratory, Bos-ton, MA. Rat monoclonal antibodies to human integrinsubunit a5 (BIIG2) and cytokeratin (7D3) were made inthis laboratory and described previously.7 Mouse mono-clonal antibody to HLA-G (IgM) was also produced in thislaboratory.23 Species-specific monoclonal antibodies toIgG and IgM were obtained from Jackson ImmunoRe-search Laboratories (West Grove, PA).
Highly purified cytotrophoblasts were cultured at aconcentration of 2.5 x 105 cells/ml on Matrigel-coated(Collaborative Research, Bedford, MA) 12-mm coverslipsin serum-free medium supplemented with 2% Nutridoma.For time zero analysis, cytotrophoblasts were allowed toadhere to the coverslips for 10 minutes at 370C. After 0,12, 24, 36, and 48 hours of culture, the cells were washedwith phosphate-buffered saline (PBS) and fixed in 2.5%paraformaldehyde at room temperature for 10 minutes.Next, cells were permeabilized with 100% methanol at-200C, and nonspecific reactivity was blocked by incu-bating them with 0.2% bovine serum albumin for 20 min-utes. Primary antibody was added at 40C for 10 to 12hours. Then the cells were washed in PBS and incubatedwith species-specific secondary antibodies at room tem-perature for 1 hour. In all cases, the cells were doublestained with antibodies that detect cytokeratin as de-scribed previously.8 Cytokeratin-positive cells werecounted on an Axiophot microscope (Zeiss), and thepercentage of these cells that expressed integrin al ora5 and HLA-G was calculated.
ImmunoblottingConditioned medium was collected from control and pre-eclamptic cytotrophoblasts at 12-hour intervals for 3 days
and stored at -800C. MMP-9 protein in these sampleswas detected by immunoblotting as previously de-scribed.10 Briefly, conditioned medium (10 ,l) was solu-bilized in loading buffer, boiled for 5 minutes, subjectedto sodium dodecyl sulfate (SDS)-polyacrylamide gelelectrophoresis (10% gels) and transferred by blotting tonitrocellulose. Nonspecific reactivity was blocked by in-cubating the transfers for 1 hour in Tris-buffered salinecontaining Tween-20 (TBST; 0.01 mol/L Tris/HCI, 0.15mol/L NaCI, and 0.05% Tween-20, pH 8.0) and 5% nonfatdried milk (Carnation). The transfers were then incubatedfor 2 hours at 25°C with anti-MMP-9 antibody (7-1 1C24),which was suspended (5 ,ug/ml) in TBST containing 5%nonfat dried milk. The blots were washed three times (10minutes each time) in TBST. Primary antibody was de-tected by incubating the blot for 2 hours at 250C withhorseradish-peroxidase-conjugated rabbit anti-mouseIgG (Jackson ImmunoResearch Laboratories) diluted 1:5000 in TBST containing 5% nonfat dried milk. The mem-branes were washed in TBST, rinsed in PBS, and devel-oped for enhanced chemiluminescence with acommercially available kit (Amersham, Arlington Heights,IL).
RNA ExtractionRNA was extracted from cytotrophoblasts cultured at aconcentration of 1.0 x 106 cells/ml in serum-free mediumon Matrigel as described above. Total RNA was ex-tracted according to published methods.25 Briefly, 5 x106 cells per sample were homogenized in 500 ,ul ofguanidine buffer (4 mol/L guanidine isothiocyanate, 25mol/L sodium citrate, 0.5% sarcosyl), followed by theaddition of 50 ,ul of 2 mol/L sodium acetate (pH 4.0), 500,ul of water-saturated phenol and 100 ,ul of chloroform.After centrifugation, the RNA was pelleted from the aque-ous phase by the addition of 500 Al of isopropanol andreprecipitated from a solution containing 10 mmol/L Tris(pH 7.5), 1 mmol/L EDTA, and 0.5% SDS. The pellet wasthen washed with cold (40C) 70% ethanol, vacuum dried,and dissolved in sterile water. The concentration of RNAwas determined by measuring the absorbance at 260nm.
Northern BlottingTotal RNA (8 ,ug) was separated by formaldehyde-agar-ose gel electrophoresis, transferred to Nytran mem-branes (Schleicher and Schuell, Keene, NH), and ana-lyzed by Northern blot hybridization as describedpreviously.2627 The probe for human integrin al wassynthesized by random priming of full-length cDNA (thegift of Dr. E. Marcantonio, Columbia University) using[32P]CTP and the Klenow fragment of DNA polymerase 1.The probe for MMP-9 was a 384-bp PCR product near the5' end of the cDNA that has the least homology to MMP-2,28 and the probe for HLA-G was synthesized by ran-dom priming of the 450-bp Pvull fragment from the 3'untranslated region of HLA-G. Probes had a specificactivity of 2 x 109 dpm/,g. In all experiments, gels were

1812 Lim et alAJP December 1997, Vol. 151, No. 6
Figure 1. Cytotrophoblast (CTB) expression of integrin al protein dramatically decreases in syndrome of hemolysis, elevated liver enzymes, and low platelets(HELLP). Control CTBs isolated from the placenta of a patient who delivered at 27 weeks of gestation due to cervical incompetence were plated on Matrigel-coatedcoverslips, cultured in serum-free medium for 48 hours and then stained with monoclonal antibodies against integrin a 1 (A) and cytokeratin (CK; B). CTBs isolatedfrom the placenta of a patient who delivered at 28 weeks of gestation due to HELLP syndrome were cultured under the same conditions for 48 hours, after whichtheir integrin al (C) and CK expression (D) was assessed. By 48 hours in culture many of the control CTBs stained with an antibody against the al integrin subunit(A), whereas few of the CTBs isolated from the placenta of the patient with HELLP syndrome reacted with this antibody (C).
stained with acridine orange before transfer to ensureintegrity of the RNA samples and to confirm that equalamounts of RNA had been loaded onto each lane. Thefinal post-hybridization wash was carried out in 0.3 xSSC (0.45 M NaCI, 4.5 mmol/L sodium citrate) and 0.1%SDS at 650C.
Invasion AssayInvasion assays were performed using one of two meth-ods previously described.29'30 In the first method, 2.5 x105 cytotrophoblasts were cultured on Transwell inserts(6.5 mm; Costar, Cambridge, MA) containing polycar-bonate filters that had been coated with 10 ,l of Matrigel.After 72 hours of culture, the cells were stained withanti-cytokeratin antibody, and the number of cell projec-tions on the bottom side of the filter was counted using afluorescence microscope as described.29 In the secondmethod, after the 72-hour culture period, cells were fixedand processed for scanning electron microscopy.30 Thefilters were removed from the inserts, critical-point dried,sputter-coated with gold palladium, and then imaged inan ETEC Autoscan Model U-1 scanning electron micro-scope. Invasion was quantified by counting the numberof cells and processes on the underside of the filter.
Statistical AnalysisAll data were analyzed using analysis of variance withScheffe F-test for post hoc analysis (StatView, AbacusConcepts, Berkeley, CA). A P value of <0.05 was con-sidered significant.
ResultsWe compared the ability of cytotrophoblasts isolatedfrom normal and pre-eclamptic placentas to differentiatealong the invasive pathway by culturing these cells underconditions that promote this process. First, we assayedtheir expression of integrins and MMP-9, stage-specificantigens that modulate their invasiveness. The integrinsof interest were the al and a5 subunits. By 48 hours inculture, cells isolated from a control placenta (27 weeks)reacted with an antibody that recognizes the al subunit(Figure 1A). Cytotrophoblasts isolated from placentasthat were obtained from a pregnancy (28 weeks) compli-cated by pre-eclampsia all reacted with cytokeratin anti-body (Figure 1D), but almost none reacted with the an-ti-al antibody (Figure 1C). In contrast, cells isolated fromboth the control and pre-eclamptic placentas expressed
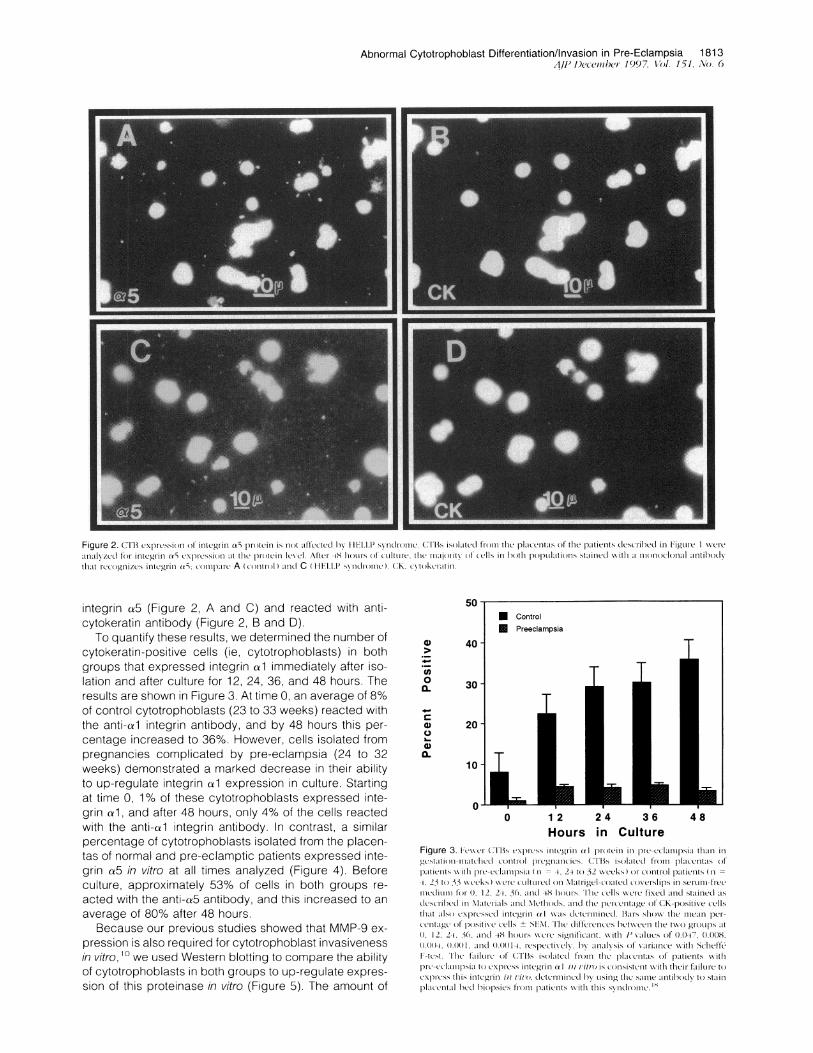
Abnormal Cytotrophoblast Differentiation/Invasion in Pre-Eclampsia 1813AtfOtv-611)e bi969) '7, Vol. 151, Ao. 6
Fiue2 ri x icso o is ' 5pocn sIo lcte yI -1,1Sn rm T Sislte r h lacla fth ptet esrbdi ig C-
'111NZeifrlitg-i Y5c pcsona h r(t_- ev tte-4 A .S dC14,1C Il 1, )1iV(I CR nh0h .)Ikto ssaie iha o o lnl_ith d
thatIccglI_c, itg-i 5 (mpr cnrl)aD C(H LPsn rmc.(K vo c li
integrin ca5 (Figure 2, A and C) and reacted with anti-cytokeratin antibody (Figure 2, B and D).
To quantify these results, we determined the number ofcytokeratin-positive cells (ie, cytotrophoblasts) in bothgroups that expressed integrin a1 immediately after iso-lation and after culture for 12, 24, 36, and 48 hours. Theresults are shown in Figure 3. At time 0, an average of 8%of control cytotrophoblasts (23 to 33 weeks) reacted withthe anti-cal integrin antibody, and by 48 hours this per-centage increased to 36%. However, cells isolated frompregnancies complicated by pre-eclampsia (24 to 32weeks) demonstrated a marked decrease in their abilityto up-regulate integrin al expression in culture. Startingat time 0, 1% of these cytotrophoblasts expressed inte-grin al, and after 48 hours, only 4% of the cells reactedwith the anti-ca1 integrin antibody. In contrast, a similarpercentage of cytotrophoblasts isolated from the placen-tas of normal and pre-eclamptic patients expressed inte-grin a5 in vitro at all times analyzed (Figure 4). Beforeculture, approximately 53% of cells in both groups re-acted with the anti-a5 antibody, and this increased to anaverage of 80% after 48 hours.
Because our previous studies showed that MMP-9 ex-pression is also required for cytotrophoblast invasivenessin vitro, 0 we used Western blotting to compare the abilityof cytotrophoblasts in both groups to up-regulate expres-sion of this proteinase in vitro (Figure 5). The amount of
50
._
._
0
0
U)0
a-0
40
30
20
101-
0 1 2 24 36 48Hours in Culture
Figure 3. iFcwXrt (CTBXs c'xpriss ii)iC'i.i(illi)a )1 pteit) ii) il C'-CcI,iilXipSia thltl) illgc'stati(i--inli.tChedCC Ontico )l poI'enci'ieCs CTl3s isolteCt t'l'O plejt'ltis (ofpa,ticilts witlh rtieclampsii (t = +, 24 to 32 XXeeks) ltr Comtr)ol pitienits (i-+ 23 to 33 weeC'ks) WX C't' C Liltt'(iel (M) MItriiel-c(ilted C(Xeirlips insert)iOi-t teItICCliLllii1()0, 12, 24, 36, inC 48 hrtos.X'lic ceXlls wre fix\ed inCh staineC -IsdcIsc Iil-t'Cd iti Mt,iteriIls inC Methti)Cds. CIn tite pe'cIttnlteagt(, CK-positiXve celXsthit iIXO ex\ptreXseCl inltegi ii) cr1 XXsiXtCeleeiinc- BitrX sXhtXX the i-ne-in pc-Ceitatig ()t clositiCll +ilSFI.'I' elitteel-tCIeC tXe tlle tX\XO gIlOLIIC)iS It(1 2 2+i 36. C111d -48 11OUICiS XXe SigtlitiC.iL WithX P \'.IiICSoe f O 047, ()1018,0)101+4 (+1+I, m1CI(11d )114e1[t-esptiX lC, Iy .iitIXXsis ()t Xviaiact' XXiti stClieftfe
te'Xt Ili'e iilCie1- o(o TIX iXis iIitCdCIli()h llietIlc itiXs of pitien)ts XXittptC'-C'Cliipii to e\Itress ilittC'giill C i)l Vilt( iS C OIiX tt' XXititltililCii tii
CXXpss ilis inltegl'ill l 'il'O tt'l-illiteCI 1WLXSill" thle S.le 1Illti)Odl) tOStXlritplachliti l hed hiopsicstiiri lpatiilts iwithitiiSStOldmi Xi

1814 Lim et al[tP 1.)ec7l) r d)() 1.IlI l. i\o (
eciatptiic patierti tO up-regulate IVIIVI-Y secretion in cuI-* Control ture.
Preeclampsia As a first step toward understanding why cytotropho-
blasts from placentas of pre-eclamptic patients fail tocorrectly modulate integrin al and MMP-9 protein ex-pression in vitro, we performed Northern hybridization tocompare levels of the corresponding mRNAs in thesecells with levels in gestation-matched control cytotropho-blasts. RNA was extracted from the cells at 0 hour, ie,immediately after isolation, or after culture for 12 and 36hours. Northern hybridization was performed using cDNAprobes that detect either integrin al mRNA (4.1 kb and5.2 to 6.5 kb) or MMP-9 mRNA (2.8 kb). At 0 hour, integrin
l1 mRNA was detected in control 26-week cytotropho-blasts, and there was a small increase in message level
o 1 2 2 4 3 6 4 8 after the cells had been cultured for 12 hours (Figure 6A).Hours in Culture In contrast, cytotrophoblasts isolated from the 27-week
nl,qr-ant_ of -q nra Pnrl_qmntir- nntiiont invnracc2arl co littla -1Figure 4. Simnilar pcrcentagexs of C113s 'plre'ssietegilil a)n protpein ill pri-eclampsipia ind in gestation-illtched colntrol piregnancies. 'th'le patients andIletliocds of anllx'iS iire decriiidil in Intlre 3, except that an antihiody to
iintc1Wi ti toa in aSXes lthe celIs intgiiitgin expiresion. thle ahilitv ofCTBs isolated froiii the placentas o)1 patiets xNxith pie-ecklillpsia to express
ilnteg'till nn inl vil/r is connsisient xx-tilt thilei exptession of this inte'grin i)i viltOdItetlillilneCIdV LISinI(g tilhe Sxamllte alltihodIx to staill placental hed hiopeosirsfpatiexlts w ith this svndrodin The dlit'letncexheixxShceteI txx Ot gipx \VeieStatistica'il\ significait onilx at timiiie = () 136)1hot nlOt thiealtteit ( IVAIaeSo' 0).22, (23 )0.43and(In .Iat 12 24, 30, and 48 houixs repectivCeixI)
MMP-9 in samples of conditioned medium from 26-weekcontrol cytotrophoblasts increased steadily during 60hours in culture. Cytotrophoblasts isolated from a 27-week pre-eclamptic patient also up-regulated MMP-9 se-cretion during 60 hours in culture. However, the amountdetected in the medium, quantified by scanning densi-tometry, was fourfold less than amounts in control me-dium throughout the culture period. We obtained essen-tially the same results in two other experiments in whichwe assayed the ability of cytotrophoblasts isolated from a28-week HELLP patient and a 29-week severely pre-
CON PE
'12h 36h 60h 12h 36h 60h
139 kD-
,84kD
Figure 5. C11B3 secIetii)oloNNINIPI-9) Icceax,es in HElI.xsilitdomePtPr-oteinxselexels in cioinclitionec iiecillicini of c.lis isxilateci ftonil the placenta11 of Ia ttlll.lpati.,nt ( Pt: 2 xxveeks)/and a contiiil i.ptient (CON; 20 xxweekxs wxrxei coni-par.,eci h\ cxieiNihIi,tting. Qciantificiioii if theit \INIP-i) xigalial tiiii ciMiilitici pic cclIlilptic Sxamlei lhx celeixitonietallait xi at 36 OcLIt xIS gax c\ \aiLcSof 3.8) + i.i31/(SENt ) aiici 1./2 + 1 texlsectixel(x/I< /.i5/). At 0/ IhOciiSthiis lciTrelice xxsaxcxn Illonic cit .iiitic ( 18ti ()5 1 I r/rxiix 1. + )2 I) <(i0i)i( I)5)liSi in mii c ixex the) cctl lixopieSciiteciCt NIPI()x.Nlteci lc inlctcondIi itiolled tielicitillifroill t il mO) IIII, ( c'ilit.SiiiecCi ncIcIiLC m ii tilisprot'iteiiiSax
P dUl IcI am ayId -(a.iid yuiL.. LIui I ILA iUZ,3t U tiU IILLI a
mRNA at either time point that the band is not visible inthe photograph. After 36 hours in culture, there was avery slight increase in ca1 mRNA expression by bothpre-eclamptic and control cells (data not shown). Analy-sis of cytotrophoblast integrin a1 mRNA levels in anothersample pair (24-week control/29-week pre-eclamptic)gave essentially the same results.A different blot of the same RNA samples shown in
Figure 6A was probed to detect MMP-9 mRNA (Figure6B). In accord with our protein data, no message wasdetected in control cells at 0 hour. By 12 hours MMP-9mRNA was found, and levels continued to increase dur-ing the 36-hour culture period. In contrast, no MMP-9mRNA was detected at either 0 or 12 hours in pre-eclamptic cytotrophoblasts. By 36 hours message waspresent, but the levels, quantified by scanning densitom-etry, were approximately 15-fold less than those found incontrol cells. These data suggest that the impaired abilityof cytotrophoblasts isolated from the placentas of pre-eclamptic patients to switch on expression of integrin aland MMP-9 in culture is due, at least in part, to a dramaticdecrease in the levels of the corresponding mRNAs.As these molecules mediate cytotrophoblast invasion
in vitro, we examined the ability of pre-eclamptic cytotro-phoblasts to invade Matrigel substrates. We comparedthe invasiveness of cells isolated from the placentas ofpatients with pre-eclampsia who delivered at 29, 30, 27,and 28 weeks to cells isolated from control patients whodelivered at 22, 24, 26, and 38 weeks. A graph summa-rizing these data is shown in Figure 7. In all cases, thecells isolated from patients with pre-eclampsia were sev-eralfold less invasive than the control cells. Overall, theseobservations suggest that, in pre-eclampsia, the invasiveability of cytotrophoblasts is dramatically decreased.
Finally, we also compared the ability of cytotropho-blasts in the two groups to modulate their expression ofstage-specific antigens that are likely to play a role inimmune interactions with maternal cells. In this case, theantigen of interest was HLA-G, an unusual major histo-compatibility complex class lb molecule expressed byfully differentiated cytotrophoblasts that have invaded theuterus."a Immediately after isolation, an average of 21%of control preterm cytotrophoblasts expressed HLA-G,
-1
.)
0._
a-0
c
G)
0~
n-4)
80 -
60
40
20
01
i nn( -r---+;--+ +- .- --. .1-+- K AK AF) n :- .1
* VV

Abnormal Cytotrophoblast Differentiation/Invasion in Pre-Eclampsia 1815AJP December 1997, Vol. 151, No. 6
A. PE CONOh 12h Oh 12h
alto -28S
MMP-18S
- 28S
-18S
Figure 6. CTB expression of integrin al and MMP-9 mRNA is reduced in pre-eclampsia. Total RNA was prepared from CTBs isolated from the placenta of a patientwith pre-eclampsia (PE; 27 weeks) and a gestationally matched control patient (CON; 26 weeks) and analyzed by Northern blotting as described in Materials andMethods. Control CTBs expressed al integrin mRNA immediately after isolation (0 h), and the level increased only slightly after 12 hours of culture (A). In contrast,CTBs isolated from the placenta of a pre-eclamptic patient expressed so little al mRNA at either 0 or 12 hours that the bands are not visible in the photograph.CTB expression of mRNA for MMP-9 was also affected by pre-eclampsia (B). As compared with control cells, the onset of expression was delayed and the amountdetected was decreased.
and at 48 hours, the number had increased to 47%(Figure 8). In contrast, an average of 3% of cytotropho-blasts isolated from pre-eclamptic patients expressedHLA-G immediately after isolation, rising only to 23% after48 hours.We further investigated whether HLA-G mRNA levels
were also reduced in pre-eclamptic cytotrophoblasts.The blot shown in Figure 4A was stripped and rehybrid-ized with a cDNA probe that corresponds to a 450-bpregion of the 3' untranslated region of HLA-G (Figure 9).Consistent with our previously published data,23 rela-tively high levels of HLA-G mRNA were detected in con-trol cytotrophoblasts immediately after isolation (0 hour),and levels did not change significantly after the cellswere cultured. In contrast, HLA-G mRNA was not de-tected in pre-eclamptic cytotrophoblasts at any time. Wealso compared levels of this message in two other pairsof pre-eclamptic and control cytotrophoblast samples: 1)28-week pre-eclamptic/40-week control and 2) 29-week
Cytotrophoblast Invasion
pre-eclamptic/26-week control. In both cases, HLA-GmRNA expression by pre-eclamptic cytotrophoblastswas less than 15% of the levels detected in control cells.
DiscussionOur results suggest that cytotrophoblast differentiationalong the invasive pathway is abnormal in pre-eclampsia.Specifically, the cells start to differentiate, as shown bytheir up-regulation of integrin a5 expression. But theirability to up-regulate expression of integrin al, MMP-9,and HLA-G, stage-specific antigens that are associatedwith later stages of the differentiation process, is im-paired. We also show that this defect has functional con-sequences; cytotrophoblasts isolated from the placentasof pre-eclamptic patients were much less invasive in vitro
0
._._Aen
m100 -
80
60-
40
0
00
0.* controlM PE
72 hrs
Hours In culture
Figure 7. Cytotrophoblast invasiveness is markedly reduced in pre-eclamp-sia. Invasion was quantified using the first assay method described in Mate-rials and Methods. The data are expressed as percentage of control invasion.The mean cell count (n = 4 separate experiments) of 639.3 + 191.6 (SEM)was used as the 100% value. The absolute value for pre-eclamptic cells was45.7 + 20.2. The difference between the two groups was statistically signif-icant (P = 0.037).
Hours in CultureFigure 8. CTB expression of HLA-G protein decreases in pre-eclampsia.CTBs isolated from placentas of patients with pre-eclampsia (n = 4, 24 to 32weeks) and control patients (n = 4, 23 to 33 weeks) were cultured onMatrigel-coated coverslips in serum-free medium for 0, 12, 24, 36, and 48hours. The cells were fixed and stained with a monoclonal antibody toHLA-G as described in Materials and Methods. The percentage of CK-positivecells that also expressed HLA-G was '500s of control levels when thepregnancy was complicated by pre-eclampsia. Except for time 0 (P = 0.054),the differences between the two groups were statistically significant (P valueof 0.01, 0.03, 0.002, and 0.03 at 12, 24, 36, and 48 hours, respectively).
.., . ,-. ___ n .......--7-77"-.I ........T
I;
7;zaC)
c
20 -

1816 Lim et alAJP December 1997, Vol. 151, No. 6
-266
HLA-GO -168
Figure 9. CTB expression of HLA-G mRNA is markedly reduced in pre-eclampsia. The blot shown in Figure 6A was stripped and rehybridized witha cDNA probe that corresponds to a 450-bp region of the 3' untranslatedregion of HLA-G. PE, pre-eclampsia; CON, control.
than gestation-matched control cells. These data (dia-grammed in Figure 10) suggest that, in pre-eclampsia,cytotrophoblasts differentiating along the invasive path-way cannot complete this process. As a result, they donot express normal levels of molecules that are requiredfor invasion (eg, integrin ca1 and MMP-9), a factor thatcould restrict invasion to the superficial portions of uterinearterioles, the placental pathology most frequently asso-ciated with pre-eclampsia. Our finding that HLA-G ex-pression is also impaired raises the interesting possibilitythat interactions between the invasive subpopulation ofcytotrophoblasts and maternal immune cells change inpre-eclampsia.
Understanding the end result of pre-eclampsia's effecton cytotrophoblast differentiation/invasion is helping us toidentify the factors that are critical to normal cytotropho-blast invasion. To date we have identified two circum-
Normal Pregnancy
A. MorphologyFETUS M H(place MO~~~~~~(tHerus
CTS bMW~~~~~~~~~~~U6
Zoel Zone/im zneiv
B. Differendtiationcra Smm CeliS _0 a" 1/HLAG -_w- azPI/MMpm9
3 I~ZoneI zon I/m ZoneIV
stances in which normal cytotrophoblasts exhibit ele-ments of the pre-eclampsia phenotype. The first is whenfirst-trimester cytotrophoblasts are cultured in an environ-ment of reduced oxygen tension. We hypothesized thatthe lack of endovascular invasion could lead to hypoxiaat the maternal-fetal interface, initiating a feedbackmechanism in which a reduction in oxygen tension furtherimpairs cytotrophoblast differentiation/invasion. To testthis idea experimentally, we compared the differentiativeand invasive capacity of normal first-trimester cytotropho-blasts cultured either under standard tissue culture con-ditions (20% 02) or in a hypoxic environment (2% 02). Wepreviously showed that cytotrophoblasts cultured in 20%02 undergo normal differentiation along the invasivepathway, ie, switch their repertoire of stage-specific an-tigens, and rapidly degrade ECMs on which they areplated. In hypoxia, the cells' integrin expression is alteredto the pattern observed in pre-eclampsia; ie, they ex-press integrin a5 but not cr1. In addition, their ability toinvade ECM is markedly reduced.29 These results sug-gest that a reduction in oxygen tension at the maternal-fetal interface can lead to abnormalities in cytotropho-blast differentiation/invasion that are similar to thoseobserved in pre-eclampsia.
The second circumstance in which normal cytotropho-blasts exhibit a pre-eclampsia phenotype is at term.Starting with the earliest-gestation samples that are rou-tinely available to us (6 weeks), we have noted a progres-sive decline in the cytotrophoblasts' differentiative andinvasive capacity as gestation progresses. With regard todifferentiation, cells isolated from term placentas have agreatly reduced ability to up-regulate expression of inte-grins cr5 and al,11 MMP-9,8 and HLA-G23 in vitro. Forexample, the percentage of normal 12- to 16-week cy-totrophoblasts that react with antibody against integrin ar1
PreecfampslaA fhnlowwIL iviovp ysvv
FE'TUS(plcenta 04,
8w~~,~~~~~abM MOTHmER
bod
zo I zoua/m zoeIVB. Dlfferentiadoncm SoC s u.asIptIl&[email protected].,
Z ! N g EwzarVFigure 10. Schema illustrating how pre-eclampsia changes cytotrophoblast invasion at the morphological and molecular levels. Comparison of the morphologyof the matemal-fetal interface in normal pregnancy (left, A) and in pre-eclampsia (right, A) shows that this syndrome is associated with abnormally shallow uterineinvasion and, in particular, with failure of CTB to penetrate the uterine arterioles. We theorize that these morphological abnormalities are the result of a defectin CTB differentiation (B). Our data suggest that in pre-eclampsia, the cells start to differentiate along the invasive pathway (as shown by their ability to expressintegrin a5) but do not complete this process (as shown by their reduced ability to express HLA-G, integrin arl, and MMP-9).
i

Abnormal Cytotrophoblast Differentiation/Invasion in Pre-Eclampsia 1817AJP December 1997, Vol. 151, No. 6
increases from 15% immediately after isolation to 60%after 48 hours in culture.11 In contrast, only 2% of termcytotrophoblasts initially react with the antibody, and thispercentage does not change during the culture period.11The percentage of integrin-al-positive cells noted in thegestation-matched controls for the experiments reportedhere (23 to 33 weeks; Figure 3) was intermediate be-tween the values for early second-trimester and termcytotrophoblasts. With regard to invasion, the cells' abilityto penetrate ECM substrates is dramatically reduced atterm.9 Thus, the placental defects that occur in pre-eclampsia could also be attributed to an acceleration ofthe normal, gestation-related changes that result in cy-totrophoblast loss of differentiative/invasive capacity atterm.
The data presented in this paper do not allow us todifferentiate unequivocally between these two possibleexplanations for why cytotrophoblast invasion is abnor-mal and placentation shallow in pre-eclampsia. Never-theless, taken together, our results to date tend to favorthe hypothesis that disease-related alterations in the dif-ferentiative/invasive capacity of cytotrophoblasts are theresult of placental hypoxia rather than an acceleration ofchanges that normally occur as pregnancy advances.The strongest evidence we have to support this theorycomes from analysis of the cells' integrin al expression ina variety of circumstances. Although term cells cannotup-regulate expression of this adhesion molecule in vitro(eg, differentiate1 1), cytotrophoblasts in tissue sections ofthe uterine wall express al at the end of normal preg-nancy but not in pre-eclampsia.17 The discordance inintegrin al expression in these two situations suggeststhat the effects of gestation-related changes and pre-eclampsia on cytotrophoblast differentiation may be sep-arable phenomena. If this is the case, then it is likely thatat least some of the phenotypic alterations that are asso-ciated with invasive cytotrophoblasts in pre-eclampsiacan be attributed to a reduction in oxygen tension at thematernal-fetal interface. Currently, we are continuing totest this hypothesis by determining whether hypoxia andpre-eclampsia have similar or different effects on a num-ber of other parameters that allow assessment of thecells' functional and differentiative capacity.
Our data also suggest that, by the time the symptomsof pre-eclampsia necessitate delivery, the deleterious ef-fects of the disease on cytotrophoblasts cannot be re-versed. The interesting question remains as to whetherthe inability of these cells to differentiate/invade is a pri-mary cause of the disease or is secondary to the patho-genesis. With regard to primary defects, transcriptionalmechanisms that regulate trophoblast differentiation inthe mouse have recently been described,3132 and it islikely that similar mechanisms play an important role inhuman placental development. Thus, it is possible thatalterations in the expression of these genes, or the reg-ulatory pathways in which they operate, could be asso-ciated with some cases of pre-eclampsia. However, thecritical role that these genes play in development sug-gests that such mutations might be relatively rare, andtheir effects might lead to abortion, rather than faultyplacentation. Thus, it seems more likely that the placental
defects that occur in pre-eclampsia are the downstreamconsequences of the pathogenesis of the disease. If thisis the case, then understanding the association betweenpre-eclampsia and the reduced capacity of cytotropho-blasts to differentiate/invade necessitates a better under-standing of the events that lie upstream of the placentalpathology. In either case, knowing the phenotype of thecytotrophoblast defect that is associated with pre-eclampsia is critical for determining whether the inabilityof the cells to differentiate/invade is the cause or theresult of this disease.
AcknowledgmentsWe thank Ms. Evangeline Leash for editing the manu-script.
References
1. Boyd J, Hamilton W: The Human Placenta. Cambridge, UK, Hefferand Sons, 1970, p 365
2. Pijnenborg R: Trophoblast invasion and placentation: morphologicalaspects. Trophoblast Res 1990, 4:33-50
3. Kurman RJ, Main CS, Chen HC: Intermediate trophoblast: a distinc-tive form of trophoblast with specific morphological, biochemical andfunctional features. Placenta 1984, 5:349-369
4. Brosens I, Robertson WB, Dixon HG: The physiological response ofthe vessels of the placental bed to normal pregnancy. J PatholBacteriol 1967, 93:569-579
5. Robertson WB, Brosens IA, Dixon HG: Placental bed vessels. Am JObstet Gynecol 1973, 117:294-295
6. Brosens IA, Robertson WB, Dixon HG: The role of the spiral arteries inthe pathogenesis of preeclampsia. Obstet Gynecol Annu 1972, 1:177-191
7. Damsky CH, Fitzgerald ML, Fisher SJ: Distribution patterns of extra-cellular matrix components and adhesion receptors are intricatelymodulated during first trimester cytotrophoblast differentiation alongthe invasive pathway, in vivo. J Clin Invest 1992, 89:210-222
8. Fisher SJ, Cui TY, Zhang L, Hartman L, Grahl K, Zhang GY, Tarpey J,Damsky CH: Adhesive and degradative properties of human placen-tal cytotrophoblast cells in vitro. J Cell Biol 1989, 109:891-902
9. Librach CL, Werb Z, Fitzgerald ML, Chiu K, Corwin NM, Esteves RA,Grobelny D, Galardy R, Damsky CH, Fisher SJ: 92-kD type IV colla-genase mediates invasion of human cytotrophoblasts. J Cell Biol1991, 113:437-449
10. Librach CL, Feigenbaum SL, Bass KE, Cui TY, Verastas N, SadovskyY, Quigley JP, French DL, Fisher SJ: lnterleukin-1 beta regulateshuman cytotrophoblast metalloproteinase activity and invasion invitro. J Biol Chem 1994, 269:17125-17131
11. Damsky CH, Librach C, Lim KH, Fitzgerald ML, McMaster MT, Janat-pour M, Zhou Y, Logan SK, Fisher SJ: Integrin switching regulatesnormal trophoblast invasion. Development 1994, 120:3657-3666
12. Chumbley G, King A, Robertson K, Holmes N, Loke YW: Resistanceof HLA-G and HLA-A2 transfectants to lysis by decidual NK cells. CellImmunol 1994, 155:312-322
13. Deniz G, Christmas SE, Brew R, Johnson PM: Phenotypic and func-tional cellular differences between human CD3- decidual and pe-ripheral blood leukocytes. J Immunol 1994, 152:4255-4261
14. Kovats S, Main EK, Librach C, Stubblebine M, Fisher SJ, DeMars R:A class antigen, HLA-G, expressed in human trophoblasts. Science1990, 248:220-223
15. Gerretsen G, Huisjes H, Elema JD: Morphological changes of thespiral arteries in the placental bed in relation to preeclampsia andfetal growth retardation. Br J Obstet Gynaecol 1981, 88:876-881
16. Khong T, De Wolf F, Robertson W, Brosens I: Inadequate maternalvascular response to placentation in pregnancies complicated bypre-eclampsia and by small-for-gestational age infants. Br J ObstetGynaecol 1986, 93:1049-1 059

1818 Lim et alAJP December 1997, Vol. 151, No. 6
17. Zhou Y, Damsky CH, Chiu K, Roberts JM, Fisher SJ: Preeclampsia isassociated with abnormal expression of adhesion molecules by in-vasive cytotrophoblasts. J Clin Invest 1993, 91:950-960
18. Berg C, Atrash H, Koonin L, Tucker M: Pregnancy-related mortality inthe United States, 1987-1990. Obstet Gynecol 1996, 88:161-167
19. American College of Obstetricians and Gynecologists: TechnicalBulletin: Hypertensive Diseases in Pregnancy. 1996, p 219
20. Redline RW, Patterson P: Pre-eclampsia is associated with an excessof proliferative immature intermediate trophoblast. Hum Pathol 1995,26:594-600
21. Chesley LC: Diagnosis of preeclampsia. Obstet Gynecol 1985, 65:423-425
22. Sibai BM: The HELLP syndrome (hemolysis, elevated liver enzymes,and low platelets): much ado about nothing? Am J Obstet Gynecol1990, 162:311-316
23. McMaster MT, Librach C, Zhou Y, Lim K-H, Janatpour MJ, DeMars R,Kovats S, Damsky C, Fisher SJ: Human placental HLA-G expressionis restricted to differentiated cytotrophoblasts. J Immunol 1995, 154:3771-3778
24. Ramos DN, Moll UM, Quigley JP, French DL: Inhibition of matrixmetalloproteinase 9 activation by a specific monoclonal antibody.Hybridoma 1993, 12:349-363
25. Chomczynski P, Sacchi N: Single-step method of RNA isolation byacid guanidinium thiocyanate-phenol-chloroform extraction. Anal Bio-chem 1987, 162:156-159
26. De S, McMaster M, Andrews G: Endotoxin induction of murine me-tallothionein gene expression. J Biol Chem 1990, 265:15267-15274
27. Lehrach H, Diamond D, Wozney J, Boedtker H: RNA molecular weightdeterminations by gel electrophoresis under denaturing conditions: acritical reexamination. Biochemistry 1977, 16:4743-4751
28. Wilhelm SM, Collier IE, Marmer BL, Eisen AZ, Grant GA, Goldberg GI:SV40-transformed human lung fibroblasts secrete a 92-kDa type IVcollagenase which is identical to that secreted by normal humanmacrophages [published erratum appears in J Biol Chem 1990,265(36):22570]. J Biol Chem 1989, 264:17213-17221
29. Genbacev 0, Joslin R, Damsky C, Polliotti B, Fisher S: Hypoxia altersearly gestation human cytotrophoblast differentiation/invasion in vitroand models the placental defects that occur in preeclampsia. J ClinInvest 1996, 97:540-550
30. Bass KE, Morrish D, Roth I, Bhardwaj D, Taylor R, Zhou Y, Fisher SJ:Human cytotrophoblast invasion is up-regulated by epidermal growthfactor: evidence that paracrine factors modify this process. Dev Biol1994, 164:550-561
31. Guillemot F, Nagy A, Auerbach A, Rossant J, Joyner AL: Essentialrole of Mash-2 in extraembryonic development. Nature 1994, 371:333-336
32. Cross JC, Flannery ML, Blanar MA, Steingrimsson E, Jenkins NA,Copeland NG, Rutter WJ, Werb Z: Hxt encodes a basic helix-loop-helix transcription factor that regulates trophoblast cell development.Development 1995, 121:2513-2523