1 research article - plantphysiol.org · 116 cultivar scarlett (ssp. vulgare). here, we report the...
TRANSCRIPT

1
Research article 1
Short title: Drought-inducible proline accumulation in barley 2
3
An ancestral allele of Pyrroline-5-carboxylate synthase1 promotes proline 4
accumulation and drought adaptation in cultivated barley 5
6
One-sentence summary: Allelic variation of Pyrroline-5-carboxylate synthase1 7
underlies sequence divergence across the promoter in the cultivated and wild barley 8
which modulates its drought-inducible transcriptional activity. 9
10
Shumaila Muzammil$1: [email protected] 11
Asis Shrestha$1: [email protected] 12
Said Dadshani1: [email protected] 13
Klaus Pillen2: [email protected] 14
Shahid Siddique3: [email protected] 15
Jens Léon1: [email protected] 16
Ali Ahmad Naz1*: [email protected] 17
1University of Bonn, Institute of Crop Science and Resource Conservation, 18
Department of Plant Breeding, Katzenburgweg 5, 53115 Bonn, Germany. 19
2Martin-Luther University Halle-Wittenberg, Halle (Saale), Institute of Agricultural and 20
Nutritional Sciences, Department of Plant Breeding, Betty-Heimann-Str. 3, 06120 21
Halle (Saale), Germany. 22
3University of Bonn, Institute of Crop Science and Resource Conservation, Molecular 23
Phytomedicine, Karlrobert-Kreiten-Straße 13, 53115 Bonn, Germany. 24
25
$Authors with equal contribution 26
*Corresponding author 27
PD Dr. Ali Ahmad Naz 28
Email: [email protected] 29
Phone: +49 228 732752, Fax: +49 228 732045 30
Plant Physiology Preview. Published on August 21, 2018, as DOI:10.1104/pp.18.00169
Copyright 2018 by the American Society of Plant Biologists
www.plantphysiol.orgon September 17, 2018 - Published by Downloaded from Copyright © 2018 American Society of Plant Biologists. All rights reserved.

2
31
Author contributions 32
A.A.N. and J.L. conceived the research idea and plans; S.M., A.S., S.D. and A.A.N. 33
carried out the phenotypic experiment and data analysis; K.P. established the S42IL 34
population and the high-resolution population S42IL-143HR for mapping and QTL 35
cloning; A.A.N., S.M., A.S. and J.L. performed QTL mapping and positional cloning; 36
S.S., A.S. performed the transient expression assays; A.A.N., A.S., S.D., S.M. and 37
S.S. wrote the manuscript. 38
39
Abstract 40
Water scarcity is a critical threat to global crop production. Here, we used the natural 41
diversity of barley (Hordeum vulgare) to dissect the genetic control of proline-42
mediated drought stress adaptation. Genetic mapping and positional cloning of a 43
major drought-inducible quantitative trait locus (QTL; QPro.S42-1H) revealed unique 44
allelic variation in pyrroline-5-carboxylate synthase (P5cs1) between the cultivated 45
cultivar Scarlett (ssp. vulgare) and the wild barley accession ISR42-8 (ssp. 46
spontaneum). The putative causative mutations were located in the promoter of 47
P5cs1 across the DNA binding motifs for abscisic acid-responsive element binding 48
transcription factors (ABFs). Introgression line (IL) S42IL-143 carrying the wild allele 49
of P5cs1 showed significant up-regulation of P5cs1 expression compared to Scarlett, 50
which was consistent with variation in proline accumulation under drought. Next, we 51
transiently expressed promoter::reporter constructs of ISR42-8 and Scarlett alleles in 52
Arabidopsis (Arabidopsis thaliana) mesophyll protoplasts. β-glucuronidase (GUS) 53
expression analysis showed a significantly higher activation of the ISR42-8 promoter 54
compared to Scarlett upon abscisic acid treatment. Notably, the ISR42-8 promoter 55
activity was impaired in protoplasts isolated from the loss-of-function 56
abf1abf2abf3abf4 quadruple mutant. A series of phenotypic evaluations 57
demonstrated that S42IL-143 maintained leaf water content and photosynthetic 58
activity longer than Scarlett under drought. These findings suggest that the ancestral 59
variant of P5cs1 has the potential for drought tolerance and understanding drought 60
physiology of barley and related crops. 61
www.plantphysiol.orgon September 17, 2018 - Published by Downloaded from Copyright © 2018 American Society of Plant Biologists. All rights reserved.

3
Introduction 62
Land plants evolved from algae approximately 500 million years ago and established 63
themselves in a terrestrial habitat (Graham, 1993). This evolution was possible as 64
plants developed multiple physiological and biochemical strategies to accomplish life 65
under water-limiting conditions. The biosynthesis of osmotic compounds of low 66
molecular weight, such as proline, glycine betaine, mannitol and organic acids, 67
seems to be an essential evolutionary event to conserve water in the terrestrial 68
habitat (Blum, 2017). The effect of these evolutionary changes can be seen as 69
unique adaptive strategies among the natural population (wild progenitors) of crop 70
plants. Therefore, the genetic dissection of diverse genetic resources is essential to 71
understand the drought physiology of crop plants as well as to develop drought-72
resilient cultivars. 73
Proline is a compatible solute and a highly soluble organic compound. It is located 74
mainly in the cytosol, chloroplasts and cytoplasmic compartments to protect cellular 75
components from dehydration injury during osmotic stress (Szabados and Savoure, 76
2010). In addition, it contributes to stabilizing sub-cellular structures (e.g. membranes 77
and proteins), scavenging free radicals and buffering cellular redox potential under 78
stress conditions (Ashraf and Foolad, 2007). Thus, proline is commonly referred to as 79
an osmolyte or osmoprotectant. As an osmolyte, it has a role in plant responses to 80
drought stress and climatic adaptation, including signal transduction, osmoregulation 81
and antioxidant systems (Kishor et al., 2005; Kesari et al., 2012). 82
Proline is synthesized primarily from glutamate by the action of the pyrroline-5-83
carboxylate synthase (P5CS) and pyrroline-5-carboxylate reductase (P5CR) 84
enzymes. In the first step, P5CS converts glutamate to glutamate-semialdehyde (Hu 85
et al., 1992), which is subsequently reduced by P5CR to proline (Szoke et al., 1992; 86
Verbruggen et al., 1993). In many plant species, there exist two conserved P5CS 87
genes, P5CS1 and P5CS2, which share approximately 82% nucleotide and 94% 88
amino acid sequence identity. However, these genes are differentially regulated at 89
the transcriptional level (Strizhov et al., 1997; Székely et al., 2008). In Arabidopsis 90
(Arabidopsis thaliana), P5CS2 modulates housekeeping proline biosynthesis in the 91
cytosol, whereas up-regulation of P5CS1 in the chloroplast results in drought-92
inducible proline accumulation. Arabidopsis P5CS1 is induced by osmotic and salt 93
stresses and activated by the abscisic acid (ABA)-dependent regulatory pathway as 94
well as H2O2-derived signals (Szabados and Savoure, 2010). Under ABA signals, 95
www.plantphysiol.orgon September 17, 2018 - Published by Downloaded from Copyright © 2018 American Society of Plant Biologists. All rights reserved.

4
ABA-responsive element (ABRE) binding proteins (AREB) or transcription factors 96
(ABFs) like ABF1, ABF2/AREB1, ABF3 and ABF4/AREB2 regulate the expression of 97
drought-inducible genes (Yoshida et al., 2015). ABFs are a group of basic leucine 98
zipper (bZIP) domain transcription factors, which were discovered using ABREs as 99
bait in yeast one-hybrid screens (Choi et al., 2000; Uno et al., 2000). It has been 100
reported that ABREs require other copies of ABREs or the combination of an ABRE 101
with one of several coupling elements (CE) across the promoter region to form the 102
ABA response complex (Shen et al., 1996). Hobo et al. (1999) showed that ABRE 103
with ACGT core and CE3 are functionally equivalent and can be considered as a 104
single class of cis-acting elements. 105
Although much is known about proline metabolism in model plants, the genetics and 106
utility are poorly understood as a trait in crops because of quantitative inheritance of 107
proline accumulation, especially under drought conditions (Kishor et al., 2005; 108
Szabados and Savoure, 2010). This scenario demands quantitative analysis of 109
drought-inducible proline accumulation among the natural population of crop plants. 110
In the present study, we employed a quantitative approach to dissect the genetic and 111
molecular regulation of proline accumulation under drought conditions in barley 112
(Hordeum vulgare). For this, we employed a set of introgression lines (ILs) 113
harbouring genome-wide marker-defined chromosomal segments of an ancestral wild 114
barley accession, ISR42-8 (ssp. spontaneum), in the background of spring barley 115
cultivar Scarlett (ssp. vulgare). Here, we report the positional isolation and molecular 116
analysis of a novel drought-inducible P5cs1 allele originated from wild barley 117
accession ISR42-8. 118
Results 119
P5cs1 allelic variation is associated with the major quantitative trait locus 120
(QTL) effect for drought-inducible proline accumulation in barley 121
Initial screening of drought-inducible proline accumulation was performed in IL 122
population designated as S42IL, which contains genome-wide wild barley ISR42-8 123
introgressions in the Scarlett background. This population exhibited a wide range of 124
variation in proline accumulation, especially under drought stress conditions 125
(Supplemental Fig. S1). For mapping, we compared individual ILs with the recurrent 126
parent Scarlett using the Dunnett-test. This analysis identified a major drought-127
inducible QTL QPro.S42-1H on chromosome 1H, where two ILs, S42IL-143 and 128
www.plantphysiol.orgon September 17, 2018 - Published by Downloaded from Copyright © 2018 American Society of Plant Biologists. All rights reserved.

5
S42IL-141, sharing a common wild barley introgression resulted in significantly higher 129
proline accumulation (Fig. 1A). In the next step, we confirmed this QTL effect by re-130
phenotyping the four selected ILs carrying wild and cultivated alleles at the target 131
QTL region. Quantification of these ILs along with parental genotypes showed a 132
significant increase in proline accumulation in S42IL-143 and S42IL-141 up to 2420 133
μg/g under drought stress (Fig. 1B). High concentrations of proline produce a red 134
colour after reaction with ninhydrin. Thus, a dark red colour indicated more proline in 135
the QTL allele harbouring IL, S42IL-143 and S42IL-141 (Fig. 1C). Using a diagnostic 136
marker (M-L), we confirmed that both ILs carried wild introgressions at the QPro.S42-137
1H region (Supplemental Fig. S2). Further, we validated this QTL effect and its 138
segregation using 237 BC4S2 plants derived from S42IL-143 (Supplemental Fig. S3). 139
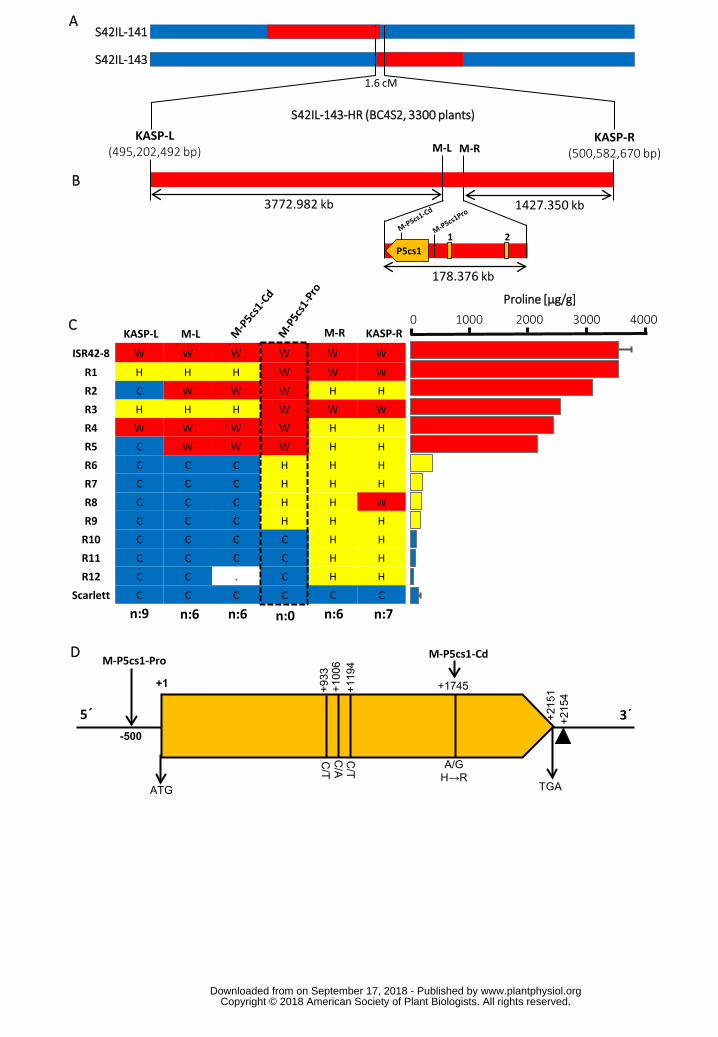
Positional cloning of QPro.S42-1H was conducted using 3300 BC4S2 plants derived 140
from S42IL-143. Initial mapping helped us to refine the targeted interval to 1.6 cM 141
between left and right border KASP markers derived from SNP:TP59951 and 142
SNP:TP3687 (Fig. 2A, Supplemental Table S1). By KASP genotyping we identified 143
85 recombinants out of 3300 plants which segregated for the targeted QTL region. 144
Later, these recombinants were quantified for proline accumulation under drought 145
(Supplemental Fig. S4). Next, we incorporated two additional markers at the left (M-146
L) and right (M-R) border of the most promising candidate gene, P5cs1, which 147
encodes a P5CS enzyme protein. By incorporating M-L and M-R, we established an 148
interval that carried no additional high confidence genes except P5cs1 (Fig. 2B, 149
Supplemental Table S2). The genotyping of M-L and M-R markers in 12 segregants 150
and their comparison with proline accumulation data revealed six recombinants at 151
each left and right border indicating that the causal mutation influencing QPro.S42-152
1H lies within P5cs1. P5cs1 was oriented in the reverse strand and comprised of 20 153
exons encoding 716 amino acids. Notably, we found two high-proline recombinants 154
(R1, R3), which were heterozygous at the 3´ UTR of P5cs1 but carried ISR42-8 155
homozygous alleles at the 5´ promoter of P5cs1. Next, we developed an additional 156
marker, M-P5cs1-Pro, at the P5cs1 promoter region (Supplemental Table S2). 157
Notably, at this marker, ISR42-8 and Scarlett alleles revealed 100% co-segregation 158
with high and low proline phenotypes, respectively. The heterozygous recombinants 159
exhibited a marginal increase in proline accumulation under drought stress conditions 160
(Fig. 2C). Sequencing of P5cs1 in ISR42-8 and Scarlett revealed a single nucleotide 161
polymorphism (SNP; A/G) in exon 15 that resulted in an amino acid substitution from 162
www.plantphysiol.orgon September 17, 2018 - Published by Downloaded from Copyright © 2018 American Society of Plant Biologists. All rights reserved.

6
histidine (ISR42-8) to arginine (Scarlett), along with three additional silent SNPs in 163
exon 7 and 9 (Fig. 2D). To prove if the amino acid substitution was associated with 164
proline variation, we developed marker M-P5cs1-Cds at the substitution site via 165
recombinant sequencing. Similar to M-L, this marker indicated the causal mutation 166
upstream of the amino acid substitution towards the 5´ promoter region of P5cs1 (Fig. 167
2C). These results suggest that the allelic variation of P5cs1 controls the drought-168
inducible QTL (QPro.S42-1H) effect in ISR42-8 and Scarlett. 169
Drought-inducible proline accumulation in S42IL-143 involves P5cs1 induction 170
Proline accumulation upon water stress is preceded by increased expression of 171
P5cs1 (Hu et al., 1992). Therefore, we hypothesized that the differential accumulation 172
of proline between Scarlett and S42IL-143 during water stress might also involve 173
changes in the expression of P5cs1. To investigate this, we analysed the expression 174
of P5cs1 in Scarlett and S42IL-143 under varying water stress conditions. The leaf 175
samples were collected at three different time points after drought stress: 3 days after 176
stress (DAS), 6 DAS and 9 DAS. RNA was extracted and analysed for the expression 177
of P5cs1 via quantitative RT-PCR (qRT-PCR). There were no significant changes in 178
the expression of P5cs1 mRNA between Scarlett and S42IL-143 under control 179
conditions. However, a significant increase in P5cs1 expression was observed in 180
S42IL-143 at 6 days after stress (DAS) and 9 DAS, whereas a significant increase 181
was only observed 9 DAS in Scarlett (Fig. 3A). Next, we tested whether this 182
significant increase in P5cs1 expression was proportional to proline accumulation. 183
For this, we quantified the proline content in the same leaf samples used for 184
expression analysis. Intriguingly, we observed that the proline accumulation in S42IL-185
143 and Scarlett followed the same trend as P5cs1 mRNA expression in response to 186
water stress (Fig. 3B). These data suggested that the QTL bearing IL S42IL-143 187
showed a drought-inducible up-regulation of P5cs1, followed by a proportional 188
increase in proline content. 189
190
Promoter divergence at the ABF binding sites putatively determines P5cs1 191
allelic variation 192
Recombination and gene expression analyses suggested that the causal mutations 193
controlling drought-inducible proline accumulation may lie in the promoter of P5cs1. 194
Therefore, we sequenced an approximately 1.5 kb region upstream of ATG in ISR42-195
8 and Scarlett. The sequence comparison revealed mutations across putative ABREs 196
www.plantphysiol.orgon September 17, 2018 - Published by Downloaded from Copyright © 2018 American Society of Plant Biologists. All rights reserved.

7
and related CE3 motifs between Scarlett and ISR42-8 (Fig. 4). Based on this, we 197
assumed that these mutations may influence the DNA binding of ABFs in the P5cs1 198
promoter. To investigate this, we established promoter::reporter constructs for ISR42-199
8 (pISR::eGFP-GUS) and Scarlett (pSCA::eGFP-GUS). Next, we transiently 200
expressed the pISR::eGFP-GUS or pSCA::eGFP-GUS constructs in protoplasts 201
isolated from Arabidopsis leaf mesophyll cells. Transfection efficiency was estimated 202
using the GFP reporter (Supplemental Fig. S5). We then incubated transfected 203
protoplasts of Col-0 ecotype in the presence of ABA (50 μM) for 4 hours and 204
analyzed the expression of GUS, ABF1, ABF2, ABF3 and ABF4 via reverse 205
transcription quantitative PCR (RT-qPCR). The expression of all four ABFs was 206
substantially induced upon ABA treatment in both pISR::eGFP-GUS- and 207
pSCA::eGFP-GUS-transfected protoplasts. Similarly, expression of the GUS gene 208
was strongly and significantly induced upon ABA treatment in pISR::eGFP-GUS 209
transfected protoplasts; however, only modest up-regulation in GUS expression was 210
observed in pSCA::eGFP-GUS-transfected protoplasts (Fig. 5A). To determine 211
whether up-regulation of GUS expression in pISR::eGFP-GUS-transfected 212
protoplasts was mediated by ABFs, we transfected this construct into mesophyll 213
protoplasts isolated from the wild-type Arabidopsis (Col-0) and loss of function 214
abf1abf2abf3abf4 quadruple mutant. Notably, we found that the up-regulation of GUS 215
activity upon ABA treatment was significantly impaired in abf1abf2abf3abf4 as 216
compared to the wild-type (Fig. 5B). Taken together, these data suggest significantly 217
different promoter activity of Scarlett and ISR42-8 alleles might be regulated in an 218
ABF-dependent manner. 219
S42IL-143 maintains higher leaf water status and photosynthetic activity under 220
drought stress compared to Scarlett 221
Plants affected by drought stress undergo changes in physiological processes 222
involved in photosynthesis, such as stomatal conductance (gs), transpiration rate (E) 223
and intercellular CO2 concentration (Ci). We investigated these parameters non-224
destructively using an infrared gas exchange analyser (LI-6400XT, LICOR Inc., 225
Lincoln, NE, USA). We found a significant reduction in gs in Scarlett under drought as 226
compared to control conditions (Supplemental Fig. S6A). S42IL-143 maintained 227
similar gs in both conditions at 3 DAS and 6 DAS, but drought reduced gs significantly 228
at 9 DAS (Supplementary Fig. S6B). The effects of drought on E were similar to 229
those of gs in both Scarlett and S42IL-143 (Supplementary Fig. S6C-D). The influx of 230
www.plantphysiol.orgon September 17, 2018 - Published by Downloaded from Copyright © 2018 American Society of Plant Biologists. All rights reserved.

8
CO2, which is essential for carbon assimilation and photosynthetic activity, is directly 231
affected by gs. We found that the internal Ci remained constant for both genotypes 232
under control conditions until 6 DAS. However, the slope of Ci decline in Scarlett was 233
40% lower than that of S42IL-143 at 9 DAS (Supplementary Fig. S6E-F). 234
Consequently, we measured the photosynthetic rate (A), which was significantly 235
reduced by drought in Scarlett at 3, 6, and 9 DAS (Fig. 6A). No significant difference 236
in A was observed in S42IL-143 at 3 and 6 DAS, but a marginal reduction due to 237
drought was observed at 9 DAS. A in S42IL-143 was 3-fold higher than in Scarlett 238
under extreme drought conditions (Fig. 6B). In addition, we measured the effective 239
quantum yield of photosystem II (YII) at steady-state photosynthesis under light using 240
a MINI-PAM-II to confirm photosynthetic activity. YII in Scarlett was significantly 241
reduced by drought at 9 DAS, whereas no significant difference was observed in 242
S42IL-143 (Fig. 6 C-D). These data suggest an increased photosynthetic activity due 243
to the QTL allele bearing-IL S42IL-143 under drought stress conditions. In addition, 244
we assessed the dynamics of the water status of Scarlett and S42IL-143 leaves 245
under control and drought stress conditions using a microwave sensor. According to 246
Dadshani et al. (2015), the microwave parameter resonant frequency shift (FRS) 247
strongly correlates with the amount of water stored in leaf tissues. Under drought, 248
Scarlett showed a significant reduction in FRS value relative to control as compared 249
to S42IL-143 (Fig. 6E). To test if these differences influenced grain yield of Scarlett 250
and S42IL-143 under drought, we performed a pot experiment in the greenhouse 251
under control and drought stress conditions. Grain weight per plant was significantly 252
lower in Scarlett compared to S42IL-143 under drought stress conditions (Fig. 6F). 253
These data indicate that drought-inducible proline accumulation has a role in 254
modulating physiological tolerance in IL S42IL-143. 255
256
Discussion 257
In the present study, we focused on the major QTL QPro.S42-1H located on 258
chromosome 1H, as it yields the strongest drought-inducible effect on proline 259
accumulation in a library of wild barley ILs. Two independent IL, S42IL-143 and 260
S42IL-141, complemented this QTL effect due to the introgression of a wild P5cs1 261
allele in the Scarlett background. S42IL-143 and S42IL-141 carried wild 262
introgressions that shared a small common segment at the P5cs1 gene locus. This 263
arrangement of introgressions was advantageous to refine the target QTL interval 264
www.plantphysiol.orgon September 17, 2018 - Published by Downloaded from Copyright © 2018 American Society of Plant Biologists. All rights reserved.

9
and to exclude the background effects of additional genes as the extent of QTL 265
QPro.S42-1H was almost similar in both ILs. Also, the near isogenic genetic 266
background of these ILs appeared more effective for the reproducibility of this 267
drought-inducible QTL during positional cloning. By this, we showed single gene 268
resolution of P5cs1 at the QTL region via high resolution recombination analysis 269
using six recombinants at each left and right borders of this gene. These findings are 270
also in line with the previous studies of proline metabolism in the model plant 271
Arabidopsis and related higher plants, which suggest a vital role of enzyme-encoding 272
gene P5CS1 for proline accumulation under drought stress conditions (Liang et al., 273
2013). 274
In the next step, we investigated sequence polymorphisms underlying genetic and 275
molecular regulation of the drought-inducible P5cs1 allele of ISR42-8. Intriguingly, we 276
found mutations in the promoters of Scarlett and ISR42-8 across the predicted 277
binding motifs of the ABF transcription factors. Previous reports have suggested that 278
ABFs are transcription factors that bind to ABREs and regulate ABA-responsive gene 279
expression (Choi et al., 2000; Uno et al., 2000). The ABF gene family is expressed in 280
vegetative tissues in response to ABA and osmotic stress in Arabidopsis, suggesting 281
a fundamental role in ABA-mediated drought stress tolerance (Fujita et al., 2011). All 282
ABF transcription factors carry four conserved domains in addition to the bZIP 283
domain (Fujita et al., 2011; Fujita et al., 2013). Transcription factors having a bZIP 284
domain target DNA duplex sites as homo- or heterodimers and bind to related but 285
distinct palindromic sequences (Ellenberger, 1994; Hurst, 1995). Shen et al. (1996) 286
shed interesting insight on the ABRE cis-elements; they found that these elements 287
require other copies of ABREs or the combination of an ABRE with one of several 288
CEs across the promoter region. These researchers also claimed that a single copy 289
of an ABRE was insufficient to activate ABA-responsive genes (Riley et al., 2008). 290
The role of multiple transcription factors binding sites (TFBS) is well documented in 291
other systems, e.g. G-box factor, in substantiating transcriptional up-regulation in 292
plants (Schulze-Lefert et al., 1989; Toniatti et al., 1990), which suggests that the 293
active role of multiple TFBS in gene up-regulation depends primarily on the targeting 294
transcription factor (TF) itself, the inter-TFB distance and the adjoining sequence of 295
the TFBS. In light of these reports, we proposed that the sequence polymorphisms in 296
ABREs and adjacent CEs across the promoter region of ISR42-8 and Scarlett may 297
influence the action of ABF transcription factors under ABA-modulated drought stress 298
www.plantphysiol.orgon September 17, 2018 - Published by Downloaded from Copyright © 2018 American Society of Plant Biologists. All rights reserved.

10
cascades. In support of this hypothesis, we performed transient expression of 299
Scarlett and ISR42-8 promoter alleles in Arabidopsis (Col-0) protoplasts with or 300
without ABA treatment. We found significantly higher GUS expression in 301
pISR::eGFP-GUS upon ABA treatment. However, a significantly impaired GUS 302
expression was observed in the protoplasts transfected with pSCA::eGFP-GUS. To 303
test the putative role of ABFs, which bind on ABRE motifs, we expressed 304
pISR::eGFP-GUS in the protoplasts of Col-0 and the abf1abf2abf3abf4 quadruple 305
mutant. Notably, GUS activity was strongly impaired in abf1abf2abf3abf4 as 306
compared to Col-0. Altogether, these data suggest that P5cs1 allelic polymorphism 307
of ABRE motifs putatively regulate differential proline accumulation in ISR42-8 and 308
Scarlett under drought stress conditions. 309
Finally, we tested whether drought-inducible proline accumulation has a role in 310
mediating drought stress tolerance in S42IL-143. Several physiological 311
measurements, such as leaf water status, -, photosynthetic parameters and 312
efficiency of photochemistry, have been widely used as markers for evaluating 313
drought stress tolerance in various plant species (Souza et al., 2004; Chaves et al., 314
2009; Liu et al., 2015). Hence, we measured these parameters in S42IL-143 and 315
Scarlett under control conditions and drought stress. Water status, measured through 316
the microwave parameter FRS, demonstrated that S42IL-143 was able to maintain 317
higher tissue water status compared to Scarlett under drought conditions. Dadshani 318
et al. (2015) also found that FRS correlates positively with water status in barley 319
leaves. Gas exchange and photosynthetic rates are considered important 320
parameters in the determination of drought stress tolerance for rain-fed agriculture. It 321
is logical that reduced gs may result in the reduction of A in plants (Lawlor and 322
Tezara, 2009; Brestic and Zivcak, 2013; Hossain et al., 2015). However, several 323
reports confirm that drought-tolerant genotypes maintain open stomata and active 324
photosynthesis, even under dehydration conditions as compared to drought-sensitive 325
genotypes (Benešová et al., 2012; Hossain et al., 2015). Our data showed significant 326
reductions in gs and A occurring early under drought stress as compared to control 327
conditions in Scarlett, whereas significant reductions under drought in S42IL-143 328
occurred only in more severe instances of drought. These parameters are direct 329
indicators to characterize the efficiency of photochemistry under varying 330
environmental conditions (Rascher et al., 2000). According to Yuan et al. (2016), 331
plants that maintain YII under drought are recognized as stress-tolerant. Our results 332
www.plantphysiol.orgon September 17, 2018 - Published by Downloaded from Copyright © 2018 American Society of Plant Biologists. All rights reserved.

11
also showed that the S42IL-143 maintained YII similar to control under drought, 333
unlike Scarlett. These data suggest that increased proline accumulation modulates 334
physiological parameters and drought tolerance in S42IL-143 due to the 335
introgression of a novel P5cs1 allele from wild barley. 336
In conclusion, the present study successfully demonstrated the isolation of a new 337
P5cs1 allele of wild origin. This QTL allele reveals promoter variation associated to 338
transcriptional induction of P5cs1, which was associated with subsequent proline 339
accumulation, thus improving drought tolerance. Future research will be focused to 340
prove a direct interaction between the causative DNA and regulatory proteins, which 341
will help to understand the molecular regulation of the ABA-driven proline cascade in 342
mediating drought tolerance. Further, we are refining the QTL allele bearing IL 343
S42IL-143 to fix the P5cs1 allele in the isogenic background of cultivated barley to 344
test the broader significance of proline accumulation for yield and yield sustainability 345
traits in barley under field conditions. 346
Materials and Methods 347
Initial screening and mapping of drought-inducible proline accumulation. 348
Initial genetic screening of proline accumulation was carried out in a population 349
(S42IL) of 72 wild barley introgression lines (IL). The S42IL lines (BC3S4:10) were 350
developed from an initial cross between the German spring cultivar Scarlett 351
(Hordeum vulgare ssp. vulgare) and the Israeli wild barley accession ISR42-8 (H. 352
vulgare ssp. spontaneum), followed by three rounds of backcrossing. The cultivar 353
Scarlett was used as the recurrent parent for subsequent backcrossing whereas 354
ISR42-8 was utilized as the donor of the drought-related traits. The development of 355
the S42IL population was described previously by Schmalenbach et al. (2008). 356
Phenotypic evaluation of the S42IL population for proline accumulation was made in 357
a split-plot design in two biological replicates of individual ILs under control and 358
drought conditions in a plastic tunnel. The treatments (control and drought) were 359
assigned to the sub-plots, within which the lines were assigned randomly. Seeds of 360
individual S42IL genotypes were sown in a plastic pot (22 × 22 × 26 cm) containing a 361
mixture of topsoil (40%) and natural sand (60%) (Terrasoil®, Cordel & Sohn, Salm, 362
Germany). Plants were watered three times a day using a drip irrigation system 363
(Netafilm, Adelaide, Australia). Echo2 sensors (Decagon Dev., Pullman WA, USA) 364
were used to determine the volumetric moisture content (VMC) digitally with the 365
frequency domain technique. The drought stress treatment was performed 30 days 366
www.plantphysiol.orgon September 17, 2018 - Published by Downloaded from Copyright © 2018 American Society of Plant Biologists. All rights reserved.

12
after sowing (DAS) by eliminating the water supply completely at plant development 367
stage BBCH 29-31 (Lancashire et al., 1991). The plants were exposed to stress for 368
26 days until the VMC reached the maximum drought stress threshold that is close to 369
the wilting point (VMC near 0%). The control block was kept under a continuous 370
supply of irrigation. The first fully developed leaf of individual genotypes in each block 371
was harvested and flash frozen in liquid nitrogen for drought and control conditions 372
and stored at -80°C until proline determination. Colorimetric method of proline 373
determination described by Bates et al., 1973 was adopted for proline content 374
measurement. Proline accumulation was quantified in µg/g of the harvested fresh 375
leaf material. 376
The S42IL population was genotyped using an Illumina 1,536 SNP-array and 377
genotyping by sequencing approaches which have been already described 378
(Schmalenbach et al., 2011; Honsdorf et al., 2014). This SNP map was compared 379
among the individual introgression lines to map the QTL region associated with 380
drought-inducible proline accumulation. ILs showing significant drought-inducible 381
proline accumulation were identified by comparing the S42IL population with the 382
recurrent parent Scarlett under control and drought stress conditions using Dunnett-383
test (Dunnett, 1955; Naz et al., 2014). 384
385
QTL validation 386
The major QTL effect QPro.S42-1H was validated by quantifying two wild allele-387
bearing ILs, S42IL-143 and S42IL-141, in comparison to two cultivated allele-bearing 388
ILs, S42IL-130 and S42IL-133, for proline accumulation under control and drought 389
stress conditions inside the growth chamber. For this, each IL was evaluated in five 390
biological replicates along with the parental genotypes. The growth chamber was 391
supplied with 12 hours of artificial light at 22±2 °C with 50-60% relative humidity. For 392
proline content measurement, two week-old seedlings were exposed to drought 393
stress by eliminating water supply completely. 394
To test the segregation of QTL alleles, the experiment was performed in the S42IL-395
143HR population derived from S42IL-143. This population was developed via 396
backcrossing of QTL bearing S42IL-143 with Scarlett and two further rounds of 397
selfing (i.e. BC4S2) as described previously (Schmalenbach et al., 2008). Seeds of 398
the S42IL-143HR population and parents were sown in climate chambers under 399
control conditions. The pots were randomized after sowing a single seed per pot (10 400
www.plantphysiol.orgon September 17, 2018 - Published by Downloaded from Copyright © 2018 American Society of Plant Biologists. All rights reserved.

13
× 10 × 12 cm). The growth chamber was supplied with 12 hours of artificial light at 401
22±2 °C with 50-60% relative humidity. Drought stress treatment was carried out 10 402
days after germination by eliminating the water supply completely. The treatment 403
pots were kept under stress, and the first fully expanded leaf was harvested from 404
drought and control conditions for proline measurement. Proline content was 405
measured nine days after drought treatment by the colorimetric procedure as 406
described previously (Bates et al., 1973). At least five independent biological 407
replicates of parents were used for proline content measurement. Higher proline 408
concentration was visualized by its reaction with ninhydrin. 409
For genotyping, a simple sequence length polymorphic (SSLP) marker M-L was 410
developed from a 3`UTR of the putative candidate gene P5cs1. This marker revealed 411
44 bp deletions in the ISR42-8 allele compared to the Scarlett allele. A total of 237 412
BC4S2 segregating progenies were genotyped using this diagnostic SSLP-marker 413
and phenotyped for proline variation under drought stress conditions. The allelic 414
polymorphism of Scarlett and ISR42-8 alleles was visualized on 2.5% standard 415
agarose gel. 416
417
Positional cloning of QTL QPro.S42-1H 418
For positional cloning, 3300 BC4S2 seeds of S42IL-143HR were sown along with 419
parental genotypes Scarlett and ISR42-8. In the next step, DNA was extracted from 420
fully expanded leaves of one-week old seedlings using the CTAB extraction method 421
according to a protocol devised by Virginia Tech Small Grains Breeding (Blacksburg, 422
Virginia, USA). For genotyping, two SNP derived KASP markers at the left (KASP-L) 423
and right (KASP-R) border of the QTL region were established. The KASP 424
genotyping was outsourced at TraitGenetics®, Gatersleben, Germany. All the 425
markers used for KASP genotyping used in this study are listed in Supplemental 426
Table S1. After KASP genotyping, recombinants were selected among the 3300 427
BC4S2 progenies that showed recombination between KASP-L and KASP-R 428
markers. These recombinants were then subjected to drought stress for 9 days, and 429
proline accumulation was measured as described previously (Bates et al., 1973). 430
Later, two additional SSLP markers, M-L and M-R, were incorporated to refine the 431
QTL region. These markers enabled refining the QTL region to a single candidate 432
gene. Later, a gene-specific marker (M-P5cs1-Pro) was established to compare the 433
co-segregation of Scarlett and ISR42-8 alleles with low and high proline phenotypes 434
www.plantphysiol.orgon September 17, 2018 - Published by Downloaded from Copyright © 2018 American Society of Plant Biologists. All rights reserved.

14
under drought stress conditions, respectively. A list of markers and corresponding 435
primers sequence information is given in Supplemental Table S2. 436
437
Expression analysis of P5cs1 mRNA 438
Expression analysis of P5cs1 mRNA was performed in Scarlett and S42IL-143 under 439
varying drought stress conditions inside the growth chamber. The experiment was 440
designed in three biological replicates for each genotype in drought stress and 441
control treatments. Drought stress treatment was introduced to 10 day-old seedlings. 442
Later, leaf samples were harvested from each plant and flash frozen in liquid nitrogen 443
and stored at -80°C until further processing. Samples were collected 3 days after 444
stress (DAS), 6 DAS and 9 DAS from both control and stress treatments. RNA 445
extraction, purification, and quantification were performed using the TRIZOL RNA 446
Isolation Protocol. The Thermo Fisher RT-qPCR kit (Thermo Fisher, Rochester, 447
USA) was used for cDNA synthesis following the manufacturer’s instructions. RT-448
qPCR was performed in 96-well plates using a 7500 fast Real-time PCR System and 449
an SYBR Green-based PCR assay using three technical replicates of each cDNA 450
sample. Each reaction contained 3 μl cDNA, 10 μl Maxima® SYBR Green/ROX 451
qPCR Master Mix, and each primer at 0.4 μM to a final volume of 20 μl. The reaction 452
mix was subjected to the following conditions: 95°C for 10 min, followed by 45 cycles 453
of 95°C for 15 s and 60°C for 30 s. Melting curves were then analysed at 95°C for 15 454
s, 60°C for 15 s, and 95°C for 15 s. In addition, a reverse transcription negative 455
control was included to assess potential genomic DNA contamination. For each 456
cDNA sample, three technical replicates were used. Elongation factor (Ef1-a) was 457
used as an internal control gene. Relative expression of P5cs1 was calculated 458
according to the 2-ΔΔCt method (Livak and Schmittgen, 2001). A list of primer 459
sequences used for expression analysis is shown in Supplemental Table S3. 460
461
Promoter analysis 462
An approximately 1.5-kb region upstream from ATG of P5cs1 was sequenced for 463
ISR42-8 and Scarlett. The promoter sequence was aligned using MAFFT alignment 464
tools (Katoh et al., 2002). DNA binding motifs across the promoter were identified 465
using MULAN analysis (Ovcharenko et al., 2005) and the plant cis-acting regulatory 466
DNA elements (PLACE) database (Higo et al., 1999) 467
468
www.plantphysiol.orgon September 17, 2018 - Published by Downloaded from Copyright © 2018 American Society of Plant Biologists. All rights reserved.

15
Transient expression of ISR42-8 and Scarlett promoter 469
Promoter regions upstream of the start codon of ISR42-8 (1534 bp) and Scarlett 470
(1537 bp) were synthesized commercially (Invitrogen, USA) and cloned in a Gateway 471
cloning vector pDONR221 (Invitrogen) according to the manufacturer’s instructions. 472
The verified promoters were fused with the eGFP-GUS tag in the expression vector 473
pBGWFS7 (Yoo et al., 2007). Transient expression analysis using Arabidopsis 474
(Arabidopsis thaliana) leaf mesophyll cells (treated with or without ABA) was 475
performed as described previously (Yoshida et al., 2002; Yoo et al., 2007; Yoshida et 476
al., 2015). A luciferase gene under the control of the 35S promoter (Karimi et al., 477
2005) was used as an internal control for GUS activity analysis between Col-0 and 478
the abf1abf2abf3abf4 quadruple mutant. The luciferase activity was performed using 479
the Luciferase Assay System (Promega, Cat. Mo. E1500) following the 480
manufacturer’s instructions. The abf1abf2abf3abf4 quadruple mutant was derived 481
from abf1 (SALK_132819), areb1/abf2 (SALK_002984), abf3 (SALK_096965) and 482
areb2/abf4 (SALK_069523) in Col-0 background (Yoshida et al., 2015). The seeds of 483
the quadruple mutant were kindly provided by Prof. Dr. Yamaguchi-Shinozaki’s lab. 484
Total RNA was extracted using an RNeasy plant mini kit (Qiagen) following the 485
manufacturer’s instructions. Contaminating DNA was digested with DNase1 using a 486
DNA-free™ DNA removal kit (Ambion), and the RNA was used to synthesize cDNA 487
using a high-capacity cDNA reverse transcription kit (Applied Biosynthesis, 488
Darmstadt, Germany) following the manufacturer’s instructions. RT-qPCR) was 489
performed with the StepOne plus real-time PCR system (Applied Biosystems) as 490
described recently (Mendy et al., 2017; Shah et al., 2017). The 18S ribosomal gene 491
was used as an endogenous control. For this, cDNA was diluted 1:100 for 18S 492
analysis. Data were analyzed using Pfaffl’s method (Pfaffl, 2001). A list of primer 493
sequences used for expression analysis is shown in Supplemental Table S4. 494
495
Determination of physiological and photosynthetic parameters 496
Various physiological parameters were analysed to determine the effect of drought 497
stress on Scarlett and the IL S42IL-143. For this purpose, a pot experiment was 498
conducted in a climate chamber with a 12h/12h light/dark photoperiod at 22±2 °C 499
with 50-60% relative humidity. The seeds of S42IL-143 were sown in 10 replications 500
each for control and treatment blocks in 1.2 L pots (10 × 10 × 12 cm) in a completely 501
randomized design. Drought stress treatment was performed at the three-leaf stage 502
www.plantphysiol.orgon September 17, 2018 - Published by Downloaded from Copyright © 2018 American Society of Plant Biologists. All rights reserved.

16
(10 days after germination) by eliminating the water supply completely. The treated 503
pots were kept under stress for 3, 6 and 9 days. For all sensor measurements, the 504
fully expanded third leaf was analysed non-destructively to detect the moisture 505
content of plant tissue (leaves) and parameters affecting photosynthetic activity. The 506
EMISENS dual mode cavity microwave resonator (EMISENS GmbH, Jülich, 507
Germany) was used to non-destructively estimate the water status of barley leaves. 508
During the assessment of a leaf with the microwave resonator, the change of the 509
resonant frequency (fr) with respect to the empty resonator was recorded. As 510
described previously by Dadshani et al. (2015), the microwave sensor parameter 511
FRS, which is the negative relative frequency shift of fr, is highly correlated with the 512
water content in tested plant material. Five measurements were taken from each 513
leaf. The infra-red gas exchange analyser (LI-6400 XT, LICOR Inc., Lincoln, NE, 514
USA) was used to measure the photosynthetic parameters, namely stomatal 515
conductance (gs), transpiration rate (E), intercellular CO2 concentration (Ci) and 516
photosynthesis rate (A). To take continuous measurements, the centre of the third 517
leaf attached to the sensor was positioned in a leaf chamber. 518
519
Grain yield evaluation of Scarlett and S42IL-143 under drought stress 520
Scarlett and S42IL-143 seeds were sown in a pot (22 x 22 x26) containing a mixture 521
of topsoil, silica sand, milled lava and peat dust (Terrasoil®, Cordel & Sohn, Salm, 522
Germany). The experiment was conducted in a randomized complete block design 523
with five replications for each genotype per treatment. Plants were watered three 524
times a day using a drip irrigation system (Netafilm, Adelaide, Australia). Two 525
drought stress treatments were applied, at tillering stage and before heading. Water 526
supply was withheld for two weeks and the pots were re-watered. Matured panicles 527
and straw were harvested and oven dried at 37°C for 72 hours. Grain weight per 528
plant was evaluated under control and drought conditions. 529
530
Accession Numbers 531
P5cs1: Gene ID (HORVU1Hr1G072780), Protein ID (A0A287G756) 532
533
Supplemental Data 534
The following supplemental materials are available. 535
www.plantphysiol.orgon September 17, 2018 - Published by Downloaded from Copyright © 2018 American Society of Plant Biologists. All rights reserved.

17
Supplemental Figure S1. Variation in proline accumulation in the S42IL population 536
under control and drought stress conditions. 537
Supplemental Figure S2. Confirmation of common wild barley introgression in 538
selected ILs. 539
Supplemental Figure S3. Segregation of QTL alleles for proline accumulation in the 540
S42IL-143-HR population. 541
Supplemental Figure S4. Variation in proline accumulation in the S42IL-143-HR 542
population under drought stress conditions. 543
Supplemental Figure S5. Transfection efficiency in Arabidopsis protoplasts. 544
Supplemental Figure S6. Gas exchange parameters of Scarlett and S42IL-143 545
under control and drought stress conditions. 546
Supplemental Table S1. Primers and sequences of KASP markers for fine mapping. 547
Supplemental Table S2. Primers used for positional cloning of P5cs1. 548
Supplemental Table S3. Primers used for expression analysis and sequencing. 549
Supplemental Table S4. Primers used for transient expression assays. 550
551
Acknowledgements 552
We thank Dr. Muhammad Ilyas for the guidelines in the protoplast transient 553
expression assay; Mrs. Karin Woitol, Mr. Süleyman Gunal and Ms. Annika Kortz for 554
providing valuable support during phenotyping and genotyping; Prof. Michael Frei, 555
Ms. Inci Vogt and Ms. Anna Backhaus for reading the manuscript. We are grateful to 556
Prof. Dr. Yamaguchi-Shinozaki for providing seeds of the abf quadruple mutant. 557
558
Funding information 559
The PhD scholarship of Mrs. Shumaila Muzammil was supported by the Higher 560
Education Commission of Pakistan. Mr. Asis Shrestha was supported by the 561
graduate college (GRK 2064) grant from the German Research Society (DFG). 562
563
Conflict of interest 564
The authors declare no conflict of interest. 565
www.plantphysiol.orgon September 17, 2018 - Published by Downloaded from Copyright © 2018 American Society of Plant Biologists. All rights reserved.

18
Figure Legends 566
Fig. 1. Genetic mapping and validation of major QTLs for proline accumulation. 567
(A) Circos plot showing the genetic map of four selected wild barley introgression 568
lines carrying ISR42-8 and Scarlett alleles at the QTL region on chromosome 1H. (B) 569
Quantification of proline QTL QPro.S42-1H in four introgression lines. Bars represent 570
the mean ± standard error (n=5). Different letters indicate significant difference of 571
genotypes and prime show differences between control and drought stress (p<0.05) 572
using Tukey’s HSD test.Pr (C) Free proline reaction with ninhydrin. A darker color 573
indicates more proline accumulation in the samples. 574
Fig. 2. Positional cloning of the major QTL QPro.S42-1H. (A) Chromosomal map 575
of wild barley introgression lines S42IL-141 and S42IL-143 overlapping at the QTL 576
region. (B) Fine mapping of the QTL region using high-resolution segregating 577
population S42IL-143HR. The refined QTL interval represents an approximately 178 578
kb region between the M-L and M-R harbouring a single high confidence gene, 579
P5cs1. The numbers 1 and 2 represent a low confidence gene and a putative 580
transposon element having no functional annotation, respectively. The physical 581
positions are according to Morex assembly HV_IBSC_PGSB_v2. (C) Comparison of 582
genotype and phenotype data among the recombinants segregating at the targeted 583
QTL interval. The cultivated, wild and heterozygous alleles are represented with 584
letters C, W and H, respectively. n=number of recombinants. Bars represent the 585
mean ± standard error (n=5) (D) Coding sequence of P5cs1 showing mutations 586
between ISR42-8 and Scarlett. Indicated SNPs represent substitutions from ISR42-8 587
to Scarlett. Black triangle represents 44 bp insertion in the 3´UTR of Scarlett. Marker 588
positions upstream and downstream of ATG are indicated by – and +, respectively. 589
Fig. 3. Expression analyses of P5cs1 and proline accumulation. (A) 590
Quantification of mRNA levels in leaves of Scarlett and S42IL-143 under control and 591
drought stress conditions via RT-qPCR. The numbers on the x-axis represent days 592
after stress (DAS). (B) Proline accumulation in leaves of Scarlett and S42IL-143 593
under control and drought stress conditions. Bars represent the mean ± standard 594
error (n=3). Asterisks indicate significance difference between control and drought 595
(*p<0.05, **p<0.01) using pair-wise t-tests. 596
Fig. 4. Promoter sequence analysis. P5cs1 promoter sequence alignment between 597
wild barley ISR42-8 and cultivar Scarlett. SNP mutations are indicated in red bold 598
letters. Core-sequence of DNA motifs are underlined with bold letters. ABRE 599
(abscisic acid-responsive element), CE3 (Coupling element 3). 600
Fig. 5. P5cs1 promoter activity analysis in Arabidopsis protoplasts upon ABA 601
treatment. (A) Relative expression of GUS, ABF1, ABF2, ABF3 and ABF4 in 602
Arabidopsis Col-0 protoplasts transfected with P5cs1 promoter::reporter constructs of 603
ISR42-8 (pISR::eGFP-GUS) and Scarlett (pSCA::eGFP-GUS). For untreated 604
samples (-ABA), data represent relative expression of the indicated genes with the 605
value of pISR::eGFP-GUS (Col-0) set to 1. For treated samples (+ABA), data 606
www.plantphysiol.orgon September 17, 2018 - Published by Downloaded from Copyright © 2018 American Society of Plant Biologists. All rights reserved.

19
represent relative expression of the indicated genes with the value of untreated 607
protoplasts set to 1. Bars represent the mean ± standard error (n=3). Asterisks 608
indicate significance difference between pISR (pISR::eGFP-GUS) and pSCA 609
(pSCA::eGFP-GUS) (**p<0.01) using pair-wise t-tests. (B) GUS activity of 610
pISR::eGFP-GUS in Arabidopsis mesophyll protoplasts of Col-0 and 611
abf1abf2abf3abf4 quadruple mutant. Bars represent the mean ± standard error (n=3). 612
Asterisks indicate significance difference between genotypes under –ABA (control) 613
and +ABA (**p<0.01, ***p<0.001) using pair-wise t-tests. 614
Fig. 6. Evaluation of physiological parameters in Scarlett and S42IL-143. (A-B) 615
Comparison of photosynthetic rate (A) in Scarlett and S42IL-143 under control and 616
different drought stress levels (0, 3, 6, 9 days after stress). (C-D) Effective quantum 617
yield of photosystem II (YII) in Scarlett and S42IL-143 under control and different 618
drought stress levels (0, 3, 6, 9 days after stress. (E) Resonance frequency shift 619
(FRS) value of Scarlett and S42IL-143 relative to control at different drought stress 620
levels (0, 3, 6, 9 days after stress). (F) Grain yield per plant in Scarlett and S42IL-143 621
under control and drought stress conditions. Data points and bars represent the 622
mean ± standard error (n=5). Asterisks indicate significance difference (*p<0.05, 623
**p<0.01) using pair-wise t-tests. 624
www.plantphysiol.orgon September 17, 2018 - Published by Downloaded from Copyright © 2018 American Society of Plant Biologists. All rights reserved.

20
References 625
Ashraf M, Foolad M (2007) Roles of glycine betaine and proline in improving plant abiotic stress 626 resistance. Environmental and Experimental Botany 59: 206-216 627
Bates L, Waldren R, Teare I (1973) Rapid determination of free proline for water-stress studies. Plant 628 and Soil 39: 205-207 629
Benešová M, Holá D, Fischer L, Jedelský PL, Hnilička F, Wilhelmová N, Rothová O, Kočová M, 630 Procházková D, Honnerová J (2012) The physiology and proteomics of drought tolerance in 631 maize: early stomatal closure as a cause of lower tolerance to short-term dehydration? PLoS 632 One 7: e38017 633
Blum A (2017) Osmotic adjustment is a prime drought stress adaptive engine in support of plant 634 production. Plant, cCell & Environment 40: 4-10 635
Brestic M, Zivcak M (2013) PSII fluorescence techniques for measurement of drought and high 636 temperature stress signal in crop plants: protocols and applications. In Molecular Stress 637 Physiology of Plants. Springer, pp87-131 638
Chaves M, Flexas J, Pinheiro C (2009) Photosynthesis under drought and salt stress: regulation 639 mechanisms from whole plant to cell. Annals of Botany 103: 551-560 640
Choi H, Hong J, Ha J, Kang J, Kim SY (2000) ABFs, a family of ABA-responsive element binding factors. 641 Journal of Biological Chemistry 275: 1723-1730 642
Dadshani S, Kurakin A, Amanov S, Hein B, Rongen H, Cranstone S, Blievernicht U, Menzel E, Léon J, 643 Klein N (2015) Non-invasive assessment of leaf water status using a dual-mode microwave 644 resonator. Plant Methods 11: 8 645
Dunnett CW (1955) A Multiple comparison procedure for comparing several treatments with a 646 control. Journal of the American Statistical Association 50: 1096-1121 647
Ellenberger T (1994) Getting a grip on DNA recognition: structures of the basic region leucine zipper, 648 and the basic region helix-loop-helix DNA-binding domains. Current Opinion in Structural 649 Biology 4: 12-21 650
Fujita Y, Fujita M, Shinozaki K, Yamaguchi-Shinozaki K (2011) ABA-mediated transcriptional 651 regulation in response to osmotic stress in plants. Journal of Plant Research 124: 509-525 652
Fujita Y, Yoshida T, Yamaguchi‐Shinozaki K (2013) Pivotal role of the AREB/ABF‐SnRK2 pathway in 653 ABRE‐mediated transcription in response to osmotic stress in plants. Physiologia Plantarum 654 147: 15-27 655
Graham LE (1993) Origin of land plants. John Wiley & Sons, Inc. 656 Higo K, Ugawa Y, Iwamoto M, Korenaga T (1999) Plant cis-acting regulatory DNA elements (PLACE) 657
database:1999. Nucleic Acids Research 27: 297-300 658 Hobo T, Asada M, Kowyama Y, Hattori T (1999) ACGT‐containing abscisic acid response element 659
(ABRE) and coupling element 3 (CE3) are functionally equivalent. The Plant Journal 19: 679-660 689 661
Honsdorf N, March TJ, Hecht A, Eglinton J, Pillen K (2014) Evaluation of juvenile drought stress 662 tolerance and genotyping by sequencing with wild barley introgression lines. Molecular 663 Breeding 34: 1475-1495 664
Hossain MM, Lam HM, Zhang J (2015) Responses in gas exchange and water status between 665 drought-tolerant and-susceptible soybean genotypes with ABA application. The Crop Journal 666 3: 500-506 667
Hu C, Delauney AJ, Verma D (1992) A bifunctional enzyme (delta 1-pyrroline-5-carboxylate 668 synthetase) catalyzes the first two steps in proline biosynthesis in plants. Proceedings of the 669 National Academy of Sciences 89: 9354-9358 670
Hurst H (1995) Transcription factors 1: bZIP proteins. Protein Profile 2. 101-168 671 Karimi M, De Meyer B, Hilson P (2005) Modular cloning and expression of tagged fluorescent protein 672
in plant cells. Trends in Plant Science 10: 103-105 673 Katoh K, Misawa K, Kuma Ki, Miyata T (2002) MAFFT: a novel method for rapid multiple sequence 674
alignment based on fast Fourier transform. Nucleic Acids Research 30: 3059-3066 675
www.plantphysiol.orgon September 17, 2018 - Published by Downloaded from Copyright © 2018 American Society of Plant Biologists. All rights reserved.

21
Kesari R, Lasky JR, Villamor JG, Des Marais DL, Chen YJ, Liu TW, Lin W, Juenger TE, Verslues PE 676 (2012) Intron-mediated alternative splicing of Arabidopsis P5CS1 and its association with 677 natural variation in proline and climate adaptation. Proceedings of the National Academy of 678 Sciences 109: 9197-9202 679
Kishor PK, Sangam S, Amrutha R, Laxmi PS, Naidu K, Rao K, Rao S, Reddy K, Theriappan P, 680 Sreenivasulu N (2005) Regulation of proline biosynthesis, degradation, uptake and transport 681 in higher plants: its implications in plant growth and abiotic stress tolerance. Current Science 682 88: 424-438 683
Lancashire PD, Bleiholder H, Boom T, Langelüddeke P, Stauss R, Weber E, Witzenberger A (1991) A 684 uniform decimal code for growth stages of crops and weeds. Annals of Applied Biology 119: 685 561-601 686
Lawlor DW, Tezara W (2009) Causes of decreased photosynthetic rate and metabolic capacity in 687 water-deficient leaf cells: a critical evaluation of mechanisms and integration of processes. 688 Annals of botany 103: 561-579 689
Liang X, Zhang L, Natarajan SK, Becker DF (2013) Proline mechanisms of stress survival. Antioxidants 690 & Redox Signaling 19: 998-1011 691
Liu Y, Zhang X, Tran H, Shan L, Kim J, Childs K, Ervin EH, Frazier T, Zhao B (2015) Assessment of 692 drought tolerance of 49 switchgrass (Panicum virgatum) genotypes using physiological and 693 morphological parameters. Biotechnology for Biofuels 8: 152 694
Livak KJ, Schmittgen TD (2001) Analysis of relative gene expression data using real-time quantitative 695 PCR and the 2− ΔΔCT method. methods 25: 402-408 696
Mendy B, Wang’ombe MW, Radakovic ZS, Holbein J, Ilyas M, Chopra D, Holton N, Zipfel C, Grundler 697 FM, Siddique S (2017) Arabidopsis leucine-rich repeat receptor–like kinase NILR1 is required 698 for induction of innate immunity to parasitic nematodes. PLoS Pathogens 13: e1006284 699
Naz AA, Arifuzzaman M, Muzammil S, Pillen K, Léon J (2014) Wild barley introgression lines revealed 700 novel QTL alleles for root and related shoot traits in the cultivated barley (Hordeum vulgare 701 L.). BMC genetics 15: 107 702
Ovcharenko I, Loots GG, Giardine BM, Hou M, Ma J, Hardison RC, Stubbs L, Miller W (2005) Mulan: 703 multiple-sequence local alignment and visualization for studying function and evolution. 704 Genome Research 15: 184-194 705
Pfaffl MW (2001) A new mathematical model for relative quantification in real-time RT–PCR. Nucleic 706 Acids Research 29: e45-e45 707
Rascher U, Liebig M, Lüttge U (2000) Evaluation of instant light‐response curves of chlorophyll 708 fluorescence parameters obtained with a portable chlorophyll fluorometer on site in the 709 field. Plant, Cell & Environment 23: 1397-1405 710
Riley T, Sontag E, Chen P, Levine A (2008) Transcriptional control of human p53-regulated genes. 711 Nature Reviews Molecular Cell Biology 9: 402-412 712
Schmalenbach I, Korber N, Pillen K (2008) Selecting a set of wild barley introgression lines and 713 verification of QTL effects for resistance to powdery mildew and leaf rust. Theoretical and 714 Applied Genetics 117: 1093-1106 715
Schmalenbach I, March TJ, Bringezu T, Waugh R, Pillen K (2011) High-resolution genotyping of wild 716 barley introgression lines and fine-mapping of the threshability locus thresh-1 using the 717 Illumina GoldenGate assay. G3: Genes, Genomes, Genetics 1: 187-196 718
Schulze-Lefert P, Dangl JL, Becker-Andre M, Hahlbrock K, Schulz W (1989) Inducible in vivo DNA 719 footprints define sequences necessary for UV light activation of the parsley chalcone 720 synthase gene. The EMBO Journal 8: 651 721
Shah SJ, Anjam MS, Mendy B, Anwer MA, Habash SS, Lozano-Torres JL, Grundler FM, Siddique S, 722 Hancock R (2017) Damage-associated responses of the host contribute to defence against 723 cyst nematodes but not root-knot nematodes. Journal of Experimental Botany 724
Shen Q, Zhang P, Ho T (1996) Modular nature of abscisic acid (ABA) response complexes: composite 725 promoter units that are necessary and sufficient for ABA induction of gene expression in 726 barley. The Plant Cell 8: 1107-1119 727
www.plantphysiol.orgon September 17, 2018 - Published by Downloaded from Copyright © 2018 American Society of Plant Biologists. All rights reserved.

22
Souza R, Machado E, Silva J, Lagôa A, Silveira J (2004) Photosynthetic gas exchange, chlorophyll 728 fluorescence and some associated metabolic changes in cowpea (Vigna unguiculata) during 729 water stress and recovery. Environmental and Experimental Botany 51: 45-56 730
Strizhov N, Ábrahám E, Ökrész L, Blickling S, Zilberstein A, Schell J, Koncz C, Szabados L (1997) 731 Differential expression of two P5CS genes controlling proline accumulation during salt‐stress 732 requires ABA and is regulated by ABA1, ABI1 and AXR2 in Arabidopsis. The Plant Journal 12: 733 557-569 734
Szabados L, Savoure A (2010) Proline: a multifunctional amino acid. Trends in Plant Science 15: 89-97 735 Székely G, Ábrahám E, Cséplő Á, Rigó G, Zsigmond L, Csiszár J, Ayaydin F, Strizhov N, Jásik J, 736
Schmelzer E (2008) Duplicated P5CS genes of Arabidopsis play distinct roles in stress 737 regulation and developmental control of proline biosynthesis. The Plant Journal 53: 11-28 738
Szoke A, Miao G-H, Hong Z, Verma DPS (1992) Subcellular location of Δ 1-pyrroline-5-carboxylate 739 reductase in root/nodule and leaf of soybean. Plant Physiology 99: 1642-1649 740
Toniatti C, Demartis A, Monaci P, Nicosia A, Ciliberto G (1990) Synergistic trans-activation of the 741 human C-reactive protein promoter by transcription factor HNF-1 binding at two distinct 742 sites. The EMBO Journal 9: 4467 743
Uno Y, Furihata T, Abe H, Yoshida R, Shinozaki K, Yamaguchi-Shinozaki K (2000) Arabidopsis basic 744 leucine zipper transcription factors involved in an abscisic acid-dependent signal transduction 745 pathway under drought and high-salinity conditions. Proceedings of the National Academy of 746 Sciences 97: 11632-11637 747
Verbruggen N, Villarroel R, Van Montagu M (1993) Osmoregulation of a pyrroline-5-carboxylate 748 reductase gene in Arabidopsis thaliana. Plant Physiology 103: 771-781 749
Yoo SD, Cho YH, Sheen J (2007) Arabidopsis mesophyll protoplasts: a versatile cell system for 750 transient gene expression analysis. Nature protocols 2: 1565-1572 751
Yoshida R, Hobo T, Ichimura K, Mizoguchi T, Takahashi F, Aronso J, Ecker JR, Shinozaki K (2002) 752 ABA-activated SnRK2 protein kinase is required for dehydration stress signaling in 753 Arabidopsis. Plant and Cell Physiology 43: 1473-1483 754
Yoshida T, Fujita Y, Maruyama K, Mogami J, Todaka D, Shinozaki K, Yamaguchi‐Shinozaki K (2015) 755 Four Arabidopsis AREB/ABF transcription factors function predominantly in gene expression 756 downstream of SnRK2 kinases in abscisic acid signalling in response to osmotic stress. Plant, 757 Cell & Environment 38: 35-49 758
Yoshida T, Fujita Y, Sayama H, Kidokoro S, Maruyama K, Mizoi J, Shinozaki K, Yamaguchi‐Shinozaki 759 K (2010) AREB1, AREB2, and ABF3 are master transcription factors that cooperatively 760 regulate ABRE‐dependent ABA signaling involved in drought stress tolerance and require ABA 761 for full activation. The Plant Journal 61: 672-685 762
Yuan X, Yang Z, Li Y, Liu Q, Han W (2016) Effects of different levels of water stress on leaf 763 photosynthetic characteristics and antioxidant enzyme activities of greenhouse tomato. 764 Photosynthetica 54: 28-39 765
766
www.plantphysiol.orgon September 17, 2018 - Published by Downloaded from Copyright © 2018 American Society of Plant Biologists. All rights reserved.

D
A B C
S42IL-143 (High proline) S42IL-141 (High proline) S42IL-130 (Low proline) S42IL-133 (Low proline)
C Stress Control
S42
IL-1
43
Sca
rlett
S42
IL-1
41
Fig. 1.
B
A
a a a
a‘ a‘ a‘ b b b
b‘ b‘ b‘
Genotype *** Treatment *** Interaction ***
www.plantphysiol.orgon September 17, 2018 - Published by Downloaded from Copyright © 2018 American Society of Plant Biologists. All rights reserved.
S42IL-143
S42IL-141
KASP-L (495,202,492 bp)
KASP-R (500,582,670 bp)
S42IL-143-HR (BC4S2, 3300 plants)
Proline [µg/g] 0 1000 2000 3000 4000 C
A
B 3772.982 kb
D
ISR42-8 W W W W W W
R1 H H H W W W R2 C W W W H H R3 H H H W W W R4 W W W W H H R5 C W W W H H R6 C C C H H H R7 C C C H H H R8 C C C H H W R9 C C C H H H
R10 C C C C H H R11 C C C C H H R12 C C . C H H
Scarlett C C C C C C
TGA ATG
A/G H→R
+1745 +1
C/T
C/A
C
/T
+100
6
+215
1
KASP-R M-R KASP-L
M-R M-L
M-L
M-P5cs1-Cd M-P5cs1-Pro
1427.350 kb
5´ 3´ -500
+215
4
n:7 n:6 n:6 n:6 n:0 n:9
+933
+119
4
178.376 kb
P5cs1
1.6 cM
1 2
www.plantphysiol.orgon September 17, 2018 - Published by Downloaded from Copyright © 2018 American Society of Plant Biologists. All rights reserved.

Fig. 3
A B
www.plantphysiol.orgon September 17, 2018 - Published by Downloaded from Copyright © 2018 American Society of Plant Biologists. All rights reserved.

CATAAGTCGTCTTTGACACGCGCCATGCGTTTGATGGGCCTCTGAGCCGCTTATCTCTCTCTCTCTTCCTTACCCTTTCCTTTCCTTATCTTTTTCCTATAAAAGCGTTGAGCCGCTCGTTC
CATAAGTCGTCTTTGACACGCGCCATGCGTTCGATGGGCCTCCGAGCCGCTTATCTCTCTCTCT--TCCTTATCCTTTCCTTTCCTTATCTTTTTCCTATAAAAGCGTTGAGCCGCTCGTTC
TTCTCCTTCTTTTTTTTCTTCGCGCGACACATCACACCCATGGGAGCACCATCCCTCATCGTCGTAGACCGGTGCACTGCCGTCGCGGCTCGTGTTGCTCCGACGTCTGCCCCTACATGTTCTCCTTCTTTTTCTTCTTCGCGCGACACATCACACCCATGGGAGCACCATCCCTCATCGTCGTAGACCGGTGCACTGCCGCCGCCGCTTGTGTTGCTCCGACGTCTGCCCCTACACG
CCACGCTCTACATCGTTGCTGCAAACCGCCGTCGCCTCGCGTCGCACCACCGCAGCTCCTGCGAAGCAGTTTGTGTCGTCGTCGCCGCGCTCCGCATCAAAGTTTTT-GCCATGGCCGCCACGCCCTACATCGTTGCTGCAAACCGCCGCCGCCTCGCGTCGCACCGCCGCAGCTCCTGCAAAGCAGTTTGTGTCGTCGTCGCCGCGCTCTGCATCAAAGTTTTTTGCCATGGCCG
TCGCATGCTGCATCCCAACATCCGCCTTCATCGACGACACTGCTGCAATGGAACGCACTGAGAGCTGCAATGGAGTGCCGCCAAGGTCGTTGGAGCTCCGTTGCGGCTCCATTGGAGCTCGCATGCTGCATCCCAACATCCGCCTTCATCGACGACACTGCTGCAATGGAACGCACCGAGAGCTGCAATGGAGTGCCGCCAAGGTCGTTGGAGCTTCGTTGCGGCTCCATTGGAGC
TTAACCAGAGCTGCAATGGAATGCCGTCGGAGTTGCAATGGAGCTTCACCGACGGCTCTAGGGAGGGGTGAGGGTGGGAGGAGCTTTTGGTGTTGCATTGGAGCTCTCGTCGTCGGCTTAACCAGAGCTGCAATGGAATGCCGTCGGAGTTGCAATGGAGCTTCACCGACGGCTCTAGGGAGGGGTGAGGGTGGGAGGAGCTTTTGGTGTTGCATTGGAGCGCTCGCCGTCGGC
TCCATTGCAGCTCCACCGGAACGATCACCGGTTACTAGACATAGCGGATGGTGTTACTATGAAGCTCTCGTCATGATTGCTCTTCGATGCCGTAATGGATGTTCGCCGGGGACTTGCGTTCCATTGCAGCTCCACCGGAACGATCACCGGTTACTAGACATAGCGTATGGTGTTACTATGAAGCTCTCGTCATGATTGCTCTTCGATGCCGTAATGGATGTTCGCCGGGGACTTGCGT
GCTCGAAATCGAAGTAGGTGGGTTGATTGCATGCACGCGAGAGGAATGCGTCATCACACATTTAAGTATCTAAGGGTCGGAAAGAATTGATCAGACGGCTAGCGACCCGATTGATTTGGGCTCGAAATCGAAGTAGGTGGGTTGATTGGATGCACGCGAGAGGAATGCGTCATCACACATTTAAGTATCTAAGGGTCGGAAAGAATTGATCAGACGGCTAGCGACCCGATTGATTTCG
CTAGAAAATCAGTCGGATGATCAATAGTAGCCGCGAGGAAAAACCGTTTTCTTGCCGTCCCTCTCCGAAGCGACGTTCGCCGGATAACATCTTGGTTCGATCGTTCGACTTGACTCAGCCTAGAAAATCAGTCGGATGATCAATAGTAGCCGCGAGGAAAAACCGTTTTCTTGCCGTCCCTCTCCGAAGCGACGTTCGCCGGATAACATCTTGGTTCGATCGTTCGACTTGACTCAGC
TCGTTCCGTGAGGCGACACACGTTTCGGTTCCCCCGTGTGAGGTGTCGTCTTTTTTTTTTTTTTTTTT----GAGATGGTGTCAGGTGTCGGCTCTAACCTGAAGCCCGAAGATGGAAGTCGTTCCGTGAGGCGACACACGTTTCGGTTCCCCCGTGTGAGGTGTCGTCTTTTTTTTTTTTTTTTTTTTTTGAGATGGTGTCAGGTGTCGACTCTAACCTGAAGCCCGAAGGTGGAAG
CAAAAAACGGACCAGACAGATATACATACACTCCAGGAGATGTACGCCACGCCGTCTCAGTATCCATATATTGATATATATCCATCTCAGAGCACGTCAGCGGCCTACCAACCCTATTGCAAAAAACGGACCAGACAGATATACATACACTCCAGGAGATGTACGCCACGCCGTCTCAGTATCCATATATTGATATATATCCATCTCAGAGCACGTCAGCGGCCTACCAACCCTATTG
TACGTGCTGTGCGTGTATCGTGCGCTCCATCTCACGAGGGGTCGATCGGACGGCCAGCGTCCCCGCGCCTCCTCCTCCGTGGAAGGCGGGTGACGCGCACGCCAACGGGACACGATACGTGCTGTGCGTGTATCGTGCGCTCCATCTCACGAGGGGTCGATCGGACGGCCAGCGTCCCCGCGCCTCCTCCTCCGTGGAAGGCGGGTGACGCGCACGCCAACGGGACACGA
CACGGGGTGGTGGTGGCGGCGGCGGCGGCGGACGCAGAGGATCACGGGGTTCGTTGGCGAGCCGCTGGGGTTTCGCTCACGGAACCAAGCACCAACCCCTCACCCCTTTAAAACCCACGGGGTGGTGGTGGCGGCGGCGGCGGCGGACGCAGAGGATCACGGGGTTCGTTGGCGAGCCGCTGGGGTTTCGCTCACGGAACCAAGCACCAACCCCTCACCCCTTTAAAACC
GCTCGGCTCTATCCCTCTCCTCTCTCTCTGACCTCCCCACACATCGCGGGTGCGGCGACGAGGGCGACGGAAAAAAAACACCGCTGATAAGAGAGAGGCGGCGCGCCGAGCCTGCCGCTCGGCTCTATCCCTCTCCTCTCTCTCTGACCTCCCCACACATCGCGGGTGCGGCGACGAGGCGACGGGAAAAAAAACACCGCAGATAAGAGAGAGGCGGCGCGCCGAGCCTGCC
CGCGGCCATGCGCGGCCATG
ISR42-8Scarlett -1542
-1420
-1301
-1183
-1065
-948
-829
-710
-591
-472
-353
-238
-123
-7
ISR42-8Scarlett
ISR42-8Scarlett
ISR42-8Scarlett
ISR42-8Scarlett
ISR42-8Scarlett
ISR42-8Scarlett
ISR42-8Scarlett
ISR42-8Scarlett
ISR42-8Scarlett
ISR42-8Scarlett
ISR42-8Scarlett
ISR42-8Scarlett
ISR42-8Scarlett
CE3
ABRE
ORF
CE3ABRE
CE3
Fig. 4
www.plantphysiol.orgon September 17, 2018 - Published by Downloaded from Copyright © 2018 American Society of Plant Biologists. All rights reserved.

Fig. 5.
A
**
B
www.plantphysiol.orgon September 17, 2018 - Published by Downloaded from Copyright © 2018 American Society of Plant Biologists. All rights reserved.

Scarlett S42IL-143
Fig. 6.
A
C
B
D
F E
A A
www.plantphysiol.orgon September 17, 2018 - Published by Downloaded from Copyright © 2018 American Society of Plant Biologists. All rights reserved.

Parsed CitationsAshraf M, Foolad M (2007) Roles of glycine betaine and proline in improving plant abiotic stress resistance. Environmental andExperimental Botany 59: 206-216
Pubmed: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
Bates L, Waldren R, Teare I (1973) Rapid determination of free proline for water-stress studies. Plant and Soil 39: 205-207Pubmed: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
Benešová M, Holá D, Fischer L, Jedelský PL, Hnilička F, Wilhelmová N, Rothová O, Kočová M, Procházková D, Honnerová J (2012) Thephysiology and proteomics of drought tolerance in maize: early stomatal closure as a cause of lower tolerance to short-termdehydration? PLoS One 7: e38017
Blum A (2017) Osmotic adjustment is a prime drought stress adaptive engine in support of plant production. Plant, cCell &Environment 40: 4-10
Pubmed: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
Brestic M, Zivcak M (2013) PSII fluorescence techniques for measurement of drought and high temperature stress signal in cropplants: protocols and applications. In Molecular Stress Physiology of Plants. Springer, pp87-131
Pubmed: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
Chaves M, Flexas J, Pinheiro C (2009) Photosynthesis under drought and salt stress: regulation mechanisms from whole plant to cell.Annals of Botany 103: 551-560
Pubmed: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
Choi H, Hong J, Ha J, Kang J, Kim SY (2000) ABFs, a family of ABA-responsive element binding factors. Journal of Biological Chemistry275: 1723-1730
Pubmed: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
Dadshani S, Kurakin A, Amanov S, Hein B, Rongen H, Cranstone S, Blievernicht U, Menzel E, Léon J, Klein N (2015) Non-invasiveassessment of leaf water status using a dual-mode microwave resonator. Plant Methods 11: 8
Pubmed: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
Dunnett CW (1955) A Multiple comparison procedure for comparing several treatments with a control. Journal of the AmericanStatistical Association 50: 1096-1121
Pubmed: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
Ellenberger T (1994) Getting a grip on DNA recognition: structures of the basic region leucine zipper, and the basic region helix-loop-helix DNA-binding domains. Current Opinion in Structural Biology 4: 12-21
Pubmed: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
Fujita Y, Fujita M, Shinozaki K, Yamaguchi-Shinozaki K (2011) ABA-mediated transcriptional regulation in response to osmotic stress inplants. Journal of Plant Research 124: 509-525
Pubmed: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
Fujita Y, Yoshida T, Yamaguchi‐Shinozaki K (2013) Pivotal role of the AREB/ABF‐SnRK2 pathway in ABRE‐mediated transcription inresponse to osmotic stress in plants. Physiologia Plantarum 147: 15-27
Pubmed: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
Graham LE (1993) Origin of land plants. John Wiley & Sons, Inc.Pubmed: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
Higo K, Ugawa Y, Iwamoto M, Korenaga T (1999) Plant cis-acting regulatory DNA elements (PLACE) database:1999. Nucleic AcidsResearch 27: 297-300
Pubmed: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
Hobo T, Asada M, Kowyama Y, Hattori T (1999) ACGT‐containing abscisic acid response element (ABRE) and coupling element 3 (CE3)are functionally equivalent. The Plant Journal 19: 679-689
Pubmed: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
www.plantphysiol.orgon September 17, 2018 - Published by Downloaded from Copyright © 2018 American Society of Plant Biologists. All rights reserved.

Honsdorf N, March TJ, Hecht A, Eglinton J, Pillen K (2014) Evaluation of juvenile drought stress tolerance and genotyping bysequencing with wild barley introgression lines. Molecular Breeding 34: 1475-1495
Pubmed: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
Hossain MM, Lam HM, Zhang J (2015) Responses in gas exchange and water status between drought-tolerant and-susceptiblesoybean genotypes with ABA application. The Crop Journal 3: 500-506
Pubmed: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
Hu C, Delauney AJ, Verma D (1992) A bifunctional enzyme (delta 1-pyrroline-5-carboxylate synthetase) catalyzes the first two steps inproline biosynthesis in plants. Proceedings of the National Academy of Sciences 89: 9354-9358
Pubmed: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
Hurst H (1995) Transcription factors 1: bZIP proteins. Protein Profile 2. 101-168Pubmed: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
Karimi M, De Meyer B, Hilson P (2005) Modular cloning and expression of tagged fluorescent protein in plant cells. Trends in PlantScience 10: 103-105
Pubmed: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
Katoh K, Misawa K, Kuma Ki, Miyata T (2002) MAFFT: a novel method for rapid multiple sequence alignment based on fast Fouriertransform. Nucleic Acids Research 30: 3059-3066
Pubmed: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
Kesari R, Lasky JR, Villamor JG, Des Marais DL, Chen YJ, Liu TW, Lin W, Juenger TE, Verslues PE (2012) Intron-mediated alternativesplicing of Arabidopsis P5CS1 and its association with natural variation in proline and climate adaptation. Proceedings of the NationalAcademy of Sciences 109: 9197-9202
Pubmed: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
Kishor PK, Sangam S, Amrutha R, Laxmi PS, Naidu K, Rao K, Rao S, Reddy K, Theriappan P, Sreenivasulu N (2005) Regulation ofproline biosynthesis, degradation, uptake and transport in higher plants: its implications in plant growth and abiotic stress tolerance.Current Science 88: 424-438
Pubmed: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
Lancashire PD, Bleiholder H, Boom T, Langelüddeke P, Stauss R, Weber E, Witzenberger A (1991) A uniform decimal code for growthstages of crops and weeds. Annals of Applied Biology 119: 561-601
Pubmed: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
Lawlor DW, Tezara W (2009) Causes of decreased photosynthetic rate and metabolic capacity in water-deficient leaf cells: a criticalevaluation of mechanisms and integration of processes. Annals of botany 103: 561-579
Pubmed: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
Liang X, Zhang L, Natarajan SK, Becker DF (2013) Proline mechanisms of stress survival. Antioxidants & Redox Signaling 19: 998-1011Pubmed: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
Liu Y, Zhang X, Tran H, Shan L, Kim J, Childs K, Ervin EH, Frazier T, Zhao B (2015) Assessment of drought tolerance of 49 switchgrass(Panicum virgatum) genotypes using physiological and morphological parameters. Biotechnology for Biofuels 8: 152
Pubmed: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
Livak KJ, Schmittgen TD (2001) Analysis of relative gene expression data using real-time quantitative PCR and the 2− ΔΔCT method.methods 25: 402-408
Pubmed: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
Mendy B, Wang'ombe MW, Radakovic ZS, Holbein J, Ilyas M, Chopra D, Holton N, Zipfel C, Grundler FM, Siddique S (2017) Arabidopsisleucine-rich repeat receptor–like kinase NILR1 is required for induction of innate immunity to parasitic nematodes. PLoS Pathogens13: e1006284
Pubmed: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
Naz AA, Arifuzzaman M, Muzammil S, Pillen K, Léon J (2014) Wild barley introgression lines revealed novel QTL alleles for root andrelated shoot traits in the cultivated barley (Hordeum vulgare L.). BMC genetics 15: 107
Pubmed: Author and TitleGoogle Scholar: Author Only Title Only Author and Title www.plantphysiol.orgon September 17, 2018 - Published by Downloaded from
Copyright © 2018 American Society of Plant Biologists. All rights reserved.

Google Scholar: Author Only Title Only Author and Title
Ovcharenko I, Loots GG, Giardine BM, Hou M, Ma J, Hardison RC, Stubbs L, Miller W (2005) Mulan: multiple-sequence local alignmentand visualization for studying function and evolution. Genome Research 15: 184-194
Pubmed: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
Pfaffl MW (2001) A new mathematical model for relative quantification in real-time RT–PCR. Nucleic Acids Research 29: e45-e45Pubmed: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
Rascher U, Liebig M, Lüttge U (2000) Evaluation of instant light‐response curves of chlorophyll fluorescence parameters obtained witha portable chlorophyll fluorometer on site in the field. Plant, Cell & Environment 23: 1397-1405
Pubmed: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
Riley T, Sontag E, Chen P, Levine A (2008) Transcriptional control of human p53-regulated genes. Nature Reviews Molecular CellBiology 9: 402-412
Pubmed: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
Schmalenbach I, Korber N, Pillen K (2008) Selecting a set of wild barley introgression lines and verification of QTL effects forresistance to powdery mildew and leaf rust. Theoretical and Applied Genetics 117: 1093-1106
Pubmed: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
Schmalenbach I, March TJ, Bringezu T, Waugh R, Pillen K (2011) High-resolution genotyping of wild barley introgression lines andfine-mapping of the threshability locus thresh-1 using the Illumina GoldenGate assay. G3: Genes, Genomes, Genetics 1: 187-196
Pubmed: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
Schulze-Lefert P, Dangl JL, Becker-Andre M, Hahlbrock K, Schulz W (1989) Inducible in vivo DNA footprints define sequencesnecessary for UV light activation of the parsley chalcone synthase gene. The EMBO Journal 8: 651
Pubmed: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
Shah SJ, Anjam MS, Mendy B, Anwer MA, Habash SS, Lozano-Torres JL, Grundler FM, Siddique S, Hancock R (2017) Damage-associated responses of the host contribute to defence against cyst nematodes but not root-knot nematodes. Journal of ExperimentalBotany
Pubmed: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
Shen Q, Zhang P, Ho T (1996) Modular nature of abscisic acid (ABA) response complexes: composite promoter units that are necessaryand sufficient for ABA induction of gene expression in barley. The Plant Cell 8: 1107-1119
Pubmed: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
Souza R, Machado E, Silva J, Lagôa A, Silveira J (2004) Photosynthetic gas exchange, chlorophyll fluorescence and some associatedmetabolic changes in cowpea (Vigna unguiculata) during water stress and recovery. Environmental and Experimental Botany 51: 45-56
Pubmed: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
Strizhov N, Ábrahám E, Ökrész L, Blickling S, Zilberstein A, Schell J, Koncz C, Szabados L (1997) Differential expression of two P5CSgenes controlling proline accumulation during salt‐stress requires ABA and is regulated by ABA1, ABI1 and AXR2 in Arabidopsis. ThePlant Journal 12: 557-569
Pubmed: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
Szabados L, Savoure A (2010) Proline: a multifunctional amino acid. Trends in Plant Science 15: 89-97Pubmed: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
Székely G, Ábrahám E, Cséplő Á, Rigó G, Zsigmond L, Csiszár J, Ayaydin F, Strizhov N, Jásik J, Schmelzer E (2008) Duplicated P5CSgenes of Arabidopsis play distinct roles in stress regulation and developmental control of proline biosynthesis. The Plant Journal 53:11-28
Pubmed: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
Szoke A, Miao G-H, Hong Z, Verma DPS (1992) Subcellular location of Δ 1-pyrroline-5-carboxylate reductase in root/nodule and leaf ofsoybean. Plant Physiology 99: 1642-1649
Pubmed: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
Toniatti C, Demartis A, Monaci P, Nicosia A, Ciliberto G (1990) Synergistic trans-activation of the human C-reactive protein promoter bytranscription factor HNF-1 binding at two distinct sites. The EMBO Journal 9: 4467
www.plantphysiol.orgon September 17, 2018 - Published by Downloaded from Copyright © 2018 American Society of Plant Biologists. All rights reserved.

Pubmed: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
Uno Y, Furihata T, Abe H, Yoshida R, Shinozaki K, Yamaguchi-Shinozaki K (2000) Arabidopsis basic leucine zipper transcription factorsinvolved in an abscisic acid-dependent signal transduction pathway under drought and high-salinity conditions. Proceedings of theNational Academy of Sciences 97: 11632-11637
Pubmed: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
Verbruggen N, Villarroel R, Van Montagu M (1993) Osmoregulation of a pyrroline-5-carboxylate reductase gene in Arabidopsis thaliana.Plant Physiology 103: 771-781
Pubmed: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
Yoo SD, Cho YH, Sheen J (2007) Arabidopsis mesophyll protoplasts: a versatile cell system for transient gene expression analysis.Nature protocols 2: 1565-1572
Pubmed: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
Yoshida R, Hobo T, Ichimura K, Mizoguchi T, Takahashi F, Aronso J, Ecker JR, Shinozaki K (2002) ABA-activated SnRK2 protein kinaseis required for dehydration stress signaling in Arabidopsis. Plant and Cell Physiology 43: 1473-1483
Pubmed: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
Yoshida T, Fujita Y, Maruyama K, Mogami J, Todaka D, Shinozaki K, Yamaguchi‐Shinozaki K (2015) Four Arabidopsis AREB/ABFtranscription factors function predominantly in gene expression downstream of SnRK2 kinases in abscisic acid signalling in responseto osmotic stress. Plant, Cell & Environment 38: 35-49
Pubmed: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
Yoshida T, Fujita Y, Sayama H, Kidokoro S, Maruyama K, Mizoi J, Shinozaki K, Yamaguchi‐Shinozaki K (2010) AREB1, AREB2, and ABF3are master transcription factors that cooperatively regulate ABRE‐dependent ABA signaling involved in drought stress tolerance andrequire ABA for full activation. The Plant Journal 61: 672-685
Pubmed: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
Yuan X, Yang Z, Li Y, Liu Q, Han W (2016) Effects of different levels of water stress on leaf photosynthetic characteristics andantioxidant enzyme activities of greenhouse tomato. Photosynthetica 54: 28-39
Pubmed: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
www.plantphysiol.orgon September 17, 2018 - Published by Downloaded from Copyright © 2018 American Society of Plant Biologists. All rights reserved.