1 copyright © 2014 elsevier inc. all rights reserved. chapter 32 neuronal network effects of drug...
TRANSCRIPT

1Copyright © 2014 Elsevier Inc. All rights reserved.
Chapter 32
Neuronal Network Effects of Drug Therapies for CNS Disorders
Carl L. Faingold

2Copyright © 2014 Elsevier Inc. All rights reserved.
FIGURE 32.1 An idealized diagram of a principal neuron in a specific nucleus within a neuronal network in the brain of an awake, behaving organism; it illustrates many of the influences that affect this “class” of neurons (N) in this nucleus. The neurons possess certain specific intrinsic properties (Φ), such as the propensity to exhibit burst firing or pacemaker activity. These cells also possess specific receptors (#) (metabotropic or ionotropic) onto which descending projections release a specific neuroactive substance. The neurons also possess ligand-gated receptors (V) (e.g. glutamate) (which have specific receptor subunits) onto which interneurons synaptically release a specific neuroactive substance that binds. Projection neurons across the midline for bilaterally connected structures release a neuroactive substance ø, which binds to its specific receptors on contralateral neurons. The principal neurons also possess the property of pH sensitivity (W). Ascending input from neurons in nuclei in the network also release a neuroactive substance (¤) that binds to its specific receptors. The neurons possess voltage-gated ion channels (X) (e.g. K+ channels) at which local ions can act. The neurons receive input from local glial cells, which release a neuroactive substance (Y) (e.g. adenosine) that acts on specific receptors for this substance. Endogenous (Endo) neuroactive agents (Z) carried via volume transmission from nearby (spillover) or distant sites via the extracellular fluid and cerebrospinal fluid to the neurons also affect the properties of these neurons. An example would be extrasynaptic GABAA receptors that respond to the low levels of “ambient” GABA in the extracellular fluid. Finally, when an exogenous (Exo) agent, such as a CNS drug, is administered, it is released via the brain blood vessels among other vectors to exert its effects on the emergent property of these principal cells to exert its therapeutic effect on a CNS disorder. Many CNS drugs are postulated to act selectively on a specific nucleus in the neuronal network for this disorder to produce the desired effect when therapeutic doses are given. The summation of all these influences determines whether the emergent property is expressed, the actual nature of that property, and its relative sensitivity in that group of neurons. This emergent property is postulated to be expressed uniquely in this network nucleus and causes this class of neurons to respond with unique sensitivity to a specific exogenous substance. Note that those influences can also be modified by brain state changes (such as sleep), which can significantly alter the emergent property, based on changes in the milieu of the principal neuron.
3Copyright © 2014 Elsevier Inc. All rights reserved.
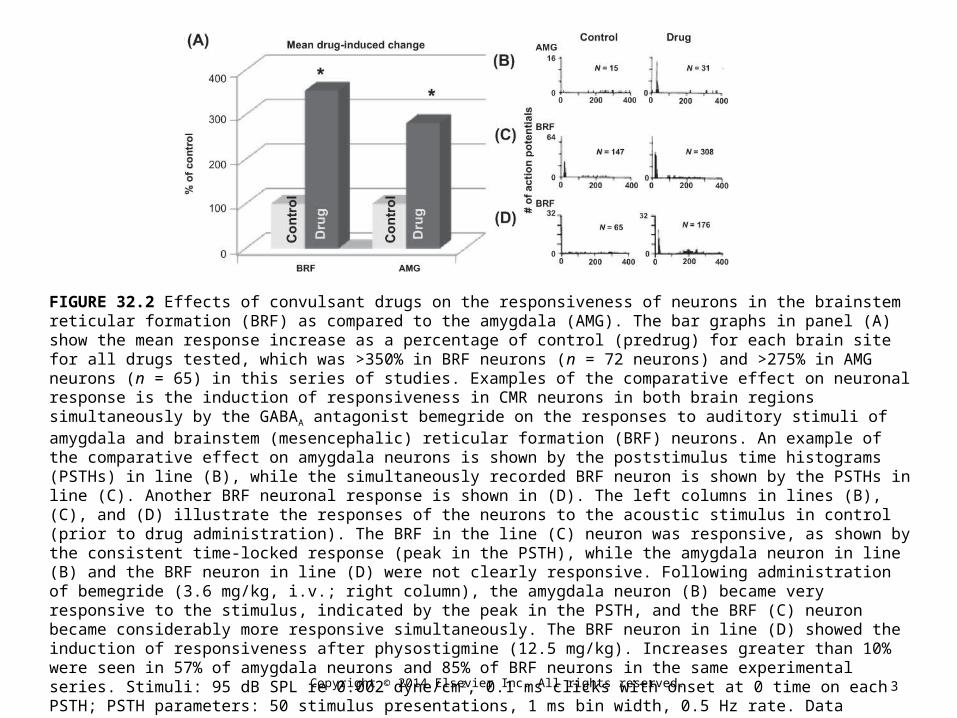
FIGURE 32.2 Effects of convulsant drugs on the responsiveness of neurons in the brainstem reticular formation (BRF) as compared to the amygdala (AMG). The bar graphs in panel (A) show the mean response increase as a percentage of control (predrug) for each brain site for all drugs tested, which was >350% in BRF neurons (n = 72 neurons) and >275% in AMG neurons (n = 65) in this series of studies. Examples of the comparative effect on neuronal response is the induction of responsiveness in CMR neurons in both brain regions simultaneously by the GABAA antagonist bemegride on the responses to auditory stimuli of amygdala and brainstem (mesencephalic) reticular formation (BRF) neurons. An example of the comparative effect on amygdala neurons is shown by the poststimulus time histograms (PSTHs) in line (B), while the simultaneously recorded BRF neuron is shown by the PSTHs in line (C). Another BRF neuronal response is shown in (D). The left columns in lines (B), (C), and (D) illustrate the responses of the neurons to the acoustic stimulus in control (prior to drug administration). The BRF in the line (C) neuron was responsive, as shown by the consistent time-locked response (peak in the PSTH), while the amygdala neuron in line (B) and the BRF neuron in line (D) were not clearly responsive. Following administration of bemegride (3.6 mg/kg, i.v.; right column), the amygdala neuron (B) became very responsive to the stimulus, indicated by the peak in the PSTH, and the BRF (C) neuron became considerably more responsive simultaneously. The BRF neuron in line (D) showed the induction of responsiveness after physostigmine (12.5 mg/kg). Increases greater than 10% were seen in 57% of amygdala neurons and 85% of BRF neurons in the same experimental series. Stimuli: 95 dB SPL re 0.002 dyne/cm2, 0.1 ms clicks with onset at 0 time on each PSTH; PSTH parameters: 50 stimulus presentations, 1 ms bin width, 0.5 Hz rate. Data obtained from unanesthetized cat with glass microelectrodes. N is the total number of action potentials in each PSTH.
(Source: Modified from Faingold et al., with permission.)

4Copyright © 2014 Elsevier Inc. All rights reserved.
FIGURE 32.3 An example of the effects of a short-acting barbiturate (thiopental) that acts to enhance GABAA receptor activation on enhanced BRF neuronal response to sensory stimuli induced by the GABAA antagonist pentylenetetrazol (PTZ). Before thiopental injection, administration of PTZ had induced extensive responsiveness to auditory and visual stimuli (A and D) in this neuron. After thiopental (5 mg/kg, i.v.), the neuron became unresponsive to both stimuli (B and E) despite continued infusion of PTZ at a rate that would normally maintain response enhancement. Responsiveness to both stimuli returned to the enhanced state 26 min after barbiturate administration (C and F). PSTH parameters: 50 stimulus presentations, 1 ms bin width, 0.5 Hz rate. Data obtained from unanesthetized cat with glass microelectrodes. N is the total number of action potentials in each PSTH.
(Source: Modified from Faingold and Caspary.)

5Copyright © 2014 Elsevier Inc. All rights reserved.
FIGURE 32.4 The action of an uncompetitive NMDA antagonist (ketamine) in a sub-anesthetic dose to reversibly block the responsiveness of neurons in a conditional multireceptive (CMR) brain region in the lateral amygdala. Panel (A) shows a bar graph of the mean effect in eight different lateral amygdala neurons in eight rats. The mean number of action potentials and poststimulus time histogram (PSTH) in control was 182.1 ± 42.0 (SEM) versus 9.6 ± 2.0 after ketamine (30 mg/kg, i.p.). Panel (B) shows a typical example of the effect of ketamine on the firing of lateral amygdala neurons. The lateral amygdala neuron exhibited an onset response before ketamine treatment [panel (B), top row]. The neuronal firing was significantly reduced (p < 0.01, paired t-test) and almost completely suppressed 15 min after ketamine treatment [panel (B), middle panel]. Four hours after ketamine treatment, the lateral amygdala neuronal response was comparable to that prior to ketamine treatment [panel (B), bottom row]. The insets [in each row of panel (B)] show examples of the digital oscilloscope tracings for each PSTH. Action potential amplitude: 300 μV. N = number of action potentials in the PSTH. Treatment was given in previously unanesthetized, awake, behaving rats with a microwire recording electrode. Acoustic stimulus parameters: 12 kHz tone burst, 100 ms duration, 5 ms rise–fall, 100 dB SPL, 0.5 Hz rate; PSTH parameters: 50 stimulus presentations, 1 ms bin width.
(Source: Based on data from Feng and Faingold,14 with permission.)

6Copyright © 2014 Elsevier Inc. All rights reserved.
FIGURE 32.5 Diagram of the neuronal network for audiogenic seizures (AGS) with possible emergent properties in each site at which systemically administered drugs that block these seizures may act at therapeutic doses. The network is organized as a hierarchy beginning with the acoustic stimulus input into the auditory pathway nuclei neurons up to the level of the inferior colliculus (IC), and then to the brainstem locomotion network nuclei to the spinal cord, to produce a sequence of neuronal activations in each nucleus during the behaviors of AGS (see Chapter 26, Fig. 26.2). Several drugs with anticonvulsant properties act on neurons in the IC, including competitive NMDA antagonists (such as 2-amino-7-phosphonoheptanoate) and a GABA uptake inhibitor (tiagabine) (note that ethanol also acts there). Other drugs that are effectively anticonvulsant exert no effect on IC neurons, including an uncompetitive NMDA antagonist (MK-801), lamotrigine, and phenytoin. Other effective anticonvulsant drugs act to reduce periaqueductal gray (PAG) neuronal firing such as phenytoin and lamotrigine, but MK-801 does not act on the PAG. Lower doses of phenytoin selectively act on neurons in the brainstem reticular formation (BRF) of the pons, but MK-801 is ineffective there. The substantia nigra reticulata (SNr) is the target of MK-801, but the effect is to increase neuronal firing. No drugs have been observed to act directly on neurons in the deep layers of the superior colliculus (DLSC). The emergent property of each nucleus is seen as a confluence of influences onto the neurons in each nucleus, including neuroactive substances (squares) released onto the neurons via synaptic transmission (ST) and from the blood vessels, and cerebrospinal and extracellular fluids via volume transmission (VT), which is also how the drugs reach each site, as shown in the diagram of an emergent property on the right of the network. CN: cochlear nucleus; SOC: superior olivary complex; NLL: nuclei of the lateral lemniscus.

7Copyright © 2014 Elsevier Inc. All rights reserved.
FIGURE 32.6 Effect of a systemically administered anticonvulsant (phenytoin) on neuronal firing in the (pontine) brainstem reticular formation (BRF) and in the periaqueductal gray (PAG) shows a relatively selective effect on the BRF in low doses that reduce seizure severity. Graph of the mean (±SEM) acoustically evoked neuronal firing as a percentage of vehicle control (open bars) on audiogenic seizures (AGS) in genetically epilepsy-prone rats (GEPR-9s) in doses of phenytoin, which significantly reduced seizure severity (low dose) or blocked seizures (high dose). Mean neuronal firing changes are shown after a phenytoin dose (mean: 6.3 mg/kg) that reduced AGS severity (i.e. blocked tonic hind limb extension) (hatched bars) and phenytoin in doses (mean: 8.3 mg/kg) that completely blocked AGS (solid bars). A significant reduction was observed in BRF neuronal firing when seizure severity was reduced (moderate seizure), which was even more greatly reduced when AGS were blocked (no seizure). However, no significant change was observed in PAG firing when the severity of AGS was reduced by phenytoin. At the higher phenytoin dose, PAG neuronal firing was also significantly reduced. These data suggest that a relatively selective effect was exerted by phenytoin on BRF neurons, as compared to the PAG, which generalized to the PAG when the dose was raised. No effect of the seizure blocking dose was seen in other neurons in the AGS network, even at the high dose. Stimulus: 12 kHz 100 ms duration tone bursts.
(Source: Based on N'Gouemo and Faingold,106 with permission.)

8Copyright © 2014 Elsevier Inc. All rights reserved.
FIGURE 32.7 An uncompetitive NMDA antagonist (dizocilpine, MK-801) blocked audiogenic seizures (AGS) in genetically epilepsy-prone rats (GEPR-9s) and in ethanol withdrawn (ETX) rats and induced significantly increased mean evoked (acoustic) neuronal responses in the substantia nigra reticulata (SNr) in vivo, as shown in (A). However, the mean effect on SNr neurons was absent in vitro (brain slices), as shown in (D). (B) An example of the effect of systemic administration of MK-801 on acoustically evoked SNr neuronal responses in a behaving epileptic rat is shown. In control, this SNr neuron exhibited a consistent response, as shown by the example in the left panel. However, 30 min after MK-801 (0.05 mg/kg, i.p.), the firing was more than doubled, as seen in the right panel, and AGS was blocked. Recovery from the effect occurred by 24 h. (D) No change was induced in SNr neurons (N = 6) in vitro. The effect of MK-801 in concentrations up to 200 μM on the mean firing of SNr neurons when depolarized was 12.8 ± 2.6 (SEM) during MK-801 perfusion, as compared to 14.2 ± 1.3 action potentials before MK-801, which was not significant. Poststimulus time histograms (PSTHs) are shown in (B), and examples of digital oscilloscope tracings of the extracellular action potentials are shown in (C). The increased firing of the SNr neuron is illustrated in the right panel. Amplitude of the action potential is 430 μV, 50 ms scan length. PSTH parameters: peak amplitude (right PSTH) axis is 32 action potentials/bin, 200 ms scan length, 1 ms bin width, 50 stimulus presentations; N = number of action potentials per PSTH.
(Source: Based on data in Faingold, with permission.)

9Copyright © 2014 Elsevier Inc. All rights reserved.
FIGURE 32.8 A network-modifying drug (gabapentin, 50 mg/kg i.p.) significantly reduced periaqueductal gray (PAG) neuronal firing and seizure severity in a repetitive epilepsy model of kindled audiogenic seizures (AGS) in GEPR-9s 60 min after administration. The same dose of this drug significantly reduced PAG neuronal firing and thermal pain in a chronic pain model (paclitaxel), but this dose had no effect on PAG neuronal firing in acute (thermal) pain. In the left pair of bars, a significantly reduced mean PAG (ventrolateral) neuronal responsiveness to amygdala (central) electrical stimulation was induced by gabapentin at all stimulus intensities tested (repeated measures ANOVA and post-hoc paired t-test). The middle set of bars shows mean thermal-evoked PAG neuronal responses in rats treated with a chronic pain-inducing protocol (paclitaxel protocol136), and a significant reduction in PAG neuronal responses was also observed 60 min after gabapentin (50 mg/kg i.p.) in normal rats. However, in an acute pain protocol (radiant heat to the paw at 53 °C), the same dose of gabapentin induced no significant change in PAG neuronal firing. Data in the left pair of bars are from awake, behaving GEPR-9s subjected to AGS kindling,134 and data in the middle and right pairs of bars are from normal rats. Significance at ∗ p < 0.01 (repeated measures ANOVA).
(Source: Based on data from Tupal and Faingold134 and Samineni et al.137)