1 caspase activity is required for engulfment of apoptotic cells
TRANSCRIPT

Caspase Activity Is Required for Engulfment of Apoptotic Cells
Boris Shklyar, Flonia Levy-Adam, Ketty Mishnaevski, Estee Kurant
Department of Anatomy and Cell Biology, The Rappaport Family Institute for Research in the Medical Sciences, Faculty of Medicine, Technion-Israel Institute ofTechnology, Haifa, Israel
Clearance of apoptotic cells by phagocytic neighbors is crucial for normal development of multicellular organisms. How-ever, how phagocytes discriminate between healthy and dying cells remains poorly understood. We focus on glial phagocy-tosis of apoptotic neurons during development of the Drosophila central nervous system. We identified phosphatidylserine(PS) as a ligand on apoptotic cells for the phagocytic receptor Six Microns Under (SIMU) and report that PS alone is notsufficient for engulfment. Our data reveal that, additionally to PS exposure, caspase activity is required for clearance ofapoptotic cells by phagocytes. Here we demonstrate that SIMU recognizes and binds PS on apoptotic cells through its N-terminal EMILIN (EMI), Nimrod 1 (NIM1), and NIM2 repeats, whereas the C-terminal NIM3 and NIM4 repeats controlSIMU affinity to PS. Based on the structure-function analysis of SIMU, we discovered a novel mechanism of internal inhi-bition responsible for differential affinities of SIMU to its ligand which might prevent elimination of living cells exposingPS on their surfaces.
The proper elimination of unwanted or aberrant cells throughapoptosis is crucial for normal development of multicellular
organisms. The final important step of apoptosis is clearance ofapoptotic cells by phagocytes. This is a complex and dynamic pro-cess, involving recruitment of phagocytes to the apoptotic cell bysecreted “find me” signals, recognition of the cell as a target forphagocytosis through “eat me” signals on the apoptotic surface,engulfment, and finally, degradation of the apoptotic particles in-side the phagosome (1–6). There are two types of phagocytes: the“professional” macrophages and immature dendritic cells and the“nonprofessional” tissue-resident neighboring cells, which are es-sential for apoptotic cell clearance during development (7–9).How phagocytes discriminate between healthy and dying cells isstill unclear. This discrimination must be highly specific and reli-able at the molecular level. Improper recognition can lead to re-moval of functional cells or survival of unwanted cells, leading tomorphogenetic defects and pathological situations.
In mammals, a large number of transmembrane receptors andsoluble bridging molecules have been shown to play a role in rec-ognition and engulfment of apoptotic particles (10–14). Many ofthese molecules recognize phosphatidylserine (PS) on the outerleaflet of apoptotic cells, raising a question of specificity and com-petition in ligand-receptor interactions. A mechanism of “tether-ing and tickling” was proposed a number of years ago (7) whichsuggests that binding of multiple phagocytic receptors to theirligands, leading to receptor clustering, is needed for engulfment ofthe apoptotic cell. The high redundancy of factors acting in themammalian systems makes it difficult to uncover the molecularand cellular mechanisms of the recognition process in vivo.
Drosophila has several receptors for apoptotic cells actingin distinct phagocytic cell populations: the CD36 homologCroquemort (CRQ) functions specifically in professional phago-cytes, the macrophages (15), whereas Draper (DRPR) and Six Mi-crons Under (SIMU) act in macrophages and also in glia duringphagocytosis of apoptotic neurons in the central nervous system(CNS) (8, 16). DRPR and SIMU belong to the recently identifiedNimrod (NIM) superfamily containing phagocytic receptors (17,18). Several members of this family have been implicated in innateimmunity (bacterial phagocytosis and apoptotic cell clearance) (8,
16, 17, 19–21) where accurate recognition of the correct targets iscrucial for efficient defense and normal development. The ligandsof most of these receptors are unknown, and elucidation of theirmechanism of action remains elusive.
The apoptotic cell death program is executed in all organismsby a specific group of cysteine proteases called caspases (22, 23).Initiator caspases start a tightly controlled proteolytic cascade toactivate themselves and effector caspases, whose activation leadsto apoptosis by cleavage of multiple cellular substrates (24, 25).Caspase activation in the fly embryo could be completely abro-gated by deletion of a genomic region containing three proapop-totic genes, reaper, grim, and hid (H99) (26).
Here we show that during Drosophila embryonic CNS devel-opment, caspase-independent PS exposure on apoptotic neuronsis not sufficient for engulfment, and additional caspase-depen-dent signals are required. We identified PS as a ligand on apoptoticcells for the phagocytic receptor SIMU. Our data demonstrate thatSIMU recognizes and binds PS on apoptotic cells through its N-ter-minal EMI, NIM1, and NIM2 repeats, whereas SIMU C-terminalNIM3 and NIM4 repeats are required for proper protein localizationand control SIMU affinity to PS. By dissecting the SIMU mode ofaction, we propose a novel internal inhibitory mechanism responsi-ble for differential affinity of a phagocytic receptor to its ligand whichis likely involved in precise uptake of appropriate targets.
MATERIALS AND METHODSFly strains and constructs. The following fly strains were obtained frompublished sources: the repoGal4 (V. Auld), UASCD8GFP (L. Luo),
Received 27 February 2013 Returned for modification 20 March 2013Accepted 4 June 2013
Published ahead of print 10 June 2013
Address correspondence to Estee Kurant, [email protected].
Supplemental material for this article may be found at http://dx.doi.org/10.1128/MCB.00233-13.
Copyright © 2013, American Society for Microbiology. All Rights Reserved.
doi:10.1128/MCB.00233-13
August 2013 Volume 33 Number 16 Molecular and Cellular Biology p. 3191–3201 mcb.asm.org 3191
Dow
nloa
ded
from
http
s://j
ourn
als.
asm
.org
/jour
nal/m
cb o
n 15
Dec
embe
r 20
21 b
y 17
7.44
.91.
41.

Df(3L)H99 (H. Steller), elavGal4 (O. Shuldiner), UASnGFP (catalog no.4775; Bloomington), and UAS-pro-dronc, UAS-ark and UASp35/TM6B(E. Arama) strains. Inducible full-length and truncated constructs of simuwere generated by cloning of the cDNA of simu in full length or withintroduced deletions into the pUASTattB vector. They also contain a C-terminal green fluorescent protein (GFP) tag. These transgenes were in-serted into the attP86 site on chromosome 3R using the QC31 system (27).All strains were raised at 25°C. w1118 flies were used as a wild-type con-trol.
SIMU protein purification and binding to apoptotic cells. The cod-ing sequence of the extracellular portion of the SIMU wild type, or of eachof the truncated constructs, was cloned into a His-Myc tag-containingvector, pCEP-Pu (28) and stably transfected into HEK293 cells, and thetagged protein was affinity purified from the conditioned medium usingTalon metal affinity beads (Clontech) according to the manufacturer’sspecifications. To induce apoptosis, S2 cells were treated with 75 �MEtoposide (Sigma) for 16 h. To evaluate protein binding to S2 cells, 6 �g ofpurified SIMU proteins was incubated with live or apoptotic S2 cells (inphosphate-buffered saline [PBS] plus 1% normal horse serum [NHS] for1.5 h). Cells were then incubated with anti-Myc antibody (Santa CruzBiotechnology) (1:100; 1 h) and fluorescent secondary antibody (AlexaFluor 488; Molecular Probes) (1:400; 40 min) and analyzed by flow cy-tometry. For each experiment, 10,000 events were collected, and the datawere analyzed using Cell Quest and FlowJo software; experiments wereperformed in triplicate and repeated three times, with similar results. Inblocking experiments, purified recombinant annexin V (BD Biosciences)was used as a phosphatidylserine (PS) masking agent. Cells were incu-bated with annexin V for 15 min before adding the SIMU proteins andprocessing as described above. In the parallel experiments, Alexa Fluor647-conjugated annexin V (Molecular Probes) was used as indicated bythe manufacturer, following incubation of the cells with purified SIMUprotein.
Phosphatidylserine binding assay. SIMU direct binding to PS wastested by a solid-phase binding assay for phospholipids (10). Polypropyl-ene plates were coated with PS, phosphatidylcholine (PC), or phosphati-dylethanolamine (PE) (Avanti Polar Lipids) at 3 �g/ml and 100 �l/well.Purified SIMU or the truncated proteins were added, and bound proteinswere detected with an anti-Myc antibody and a secondary antibody con-jugated to horseradish peroxidase (HRP).
Immunohistochemistry and imaging. Guinea pig anti-SIMU anti-body was raised against SIMU wild-type protein purified from condi-tioned medium of SIMU-transfected HEK293 cells as described in refer-ence 8. All embryos were stained by standard procedures using rabbitanti-activated caspase 3 (CM1/Abcam) (1:200). Mouse anti-GFP (1:100)is from Roche. Fluorescent secondary antibodies (Cy3 [Jackson Immuno-Research] and Alexa Fluor 488 [Molecular Probes]) were used in 1:200dilutions. All confocal images were acquired on a Zeiss LSM 700 confocalmicroscope using an EC Plan-Neofluar 40�/1.30 oil differential interfer-ence contrast (DIC) M27 objective and a Plan-Apochromat 20�/0.8 M27lens. Glycerol solution (75%) was used as the imaging medium. Imageanalysis was performed using Zeiss LSM 700 and Imaris (Bitplane) soft-ware. To quantitate the volume of apoptotic particles, confocal stacks (5sections; total, 7.5 �m) were acquired from the neural cortex of stage 16ventral nerve cords.
Live imaging was carried out by dechorionating embryos (stage 15),mounting them under halocarbon oil, and injecting 2% to 3% egg volumeAlexa Fluor 555-conjugated annexin V (Molecular Probes) or Lyso-Tracker (Molecular Probes). Recording started 30 min following injec-tion.
RESULTSPS is a ligand of SIMU on apoptotic cells. In order to understandthe mechanism underlying recognition of apoptotic particles byphagocytic receptors, we focused on the phagocytic receptorSIMU, which is required for recognition and engulfment of apop-
totic particles in vivo and binds specifically to apoptotic cells invitro (8). To identify the SIMU ligand(s) on the apoptotic surface,we took a structure-function approach. SIMU comprises a largeextracellular portion, a single transmembrane domain (TM), anda short cytoplasmic tail at the C terminus with no signaling ability(8) (Fig. 1A). The extracellular portion of the protein consists ofan N-terminal EMILIN (EMI) domain and four EGF domainsnamed Nimrod (NIM) repeats (8). To establish which domain(s)is involved in ligand recognition, we tested the ability of truncatedforms of the SIMU protein (Fig. 1A) to bind to apoptotic Drosoph-ila Schneider (S2) cells. We have previously shown that the se-creted form of SIMU (SIMU�TM) strongly binds to apoptotic S2cells in an in vitro assay based on flow cytometry (8). Here, weshow that deletion of each of the EMI, NIM1, and NIM2 domainsor of the three domains together results in no binding of SIMU toapoptotic cells (Fig. 1B). Therefore, we reasoned that EMI, NIM1,and NIM2 are required for SIMU binding to apoptotic cells. Incontrast, and to our surprise, a lack of the NIM3 domain or of theNIM4 domain or of their combination not only does not reducebut rather significantly increases SIMU binding to apoptotic cells(Fig. 1C), suggesting an inhibitory effect of these domains on EMI,NIM1, and NIM2 binding and no involvement in ligand binding.
NIM domains are characterized by a consensus sequence (Fig.S1A); however, small differences exist between NIM domains inSIMU. To test why NIM2 and NIM3 behave so differently and tounderstand whether the sequence or the position of NIM domainsdetermines their function, we performed swapping of NIM2 andNIM3 domains. We generated SIMU full-length secreted proteinscontaining two NIM2 domains (an inhibitory NIM3 domain wasreplaced by the NIM2 domain) or two NIM3 domains (the NIM2domain was replaced by the inhibitory NIM3 domain) (see Fig.S1A and B in the supplemental material). Interestingly, the pro-tein with two NIM3 domains behaved similarly to the SIMU�TMprotein, indicating that the NIM3 domain in the position of theNIM2 domain functions similarly to NIM2 in binding to PS anddoes not inhibit SIMU binding to apoptotic cells (see Fig. S1C).However, when we tested the SIMU protein containing two NIM2domains, we observed much stronger binding of the protein to S2cells, indicating that substitution of the NIM3 domain with NIM2resulted in a reduction of internal inhibition (see Fig. S1C). Weobtained the same results in the enzyme-linked immunosorbentassay (ELISA), which we performed with the same swapped pro-teins (see Fig. S1D), suggesting that the inhibitory function of theNIM3 repeat depends on its position.
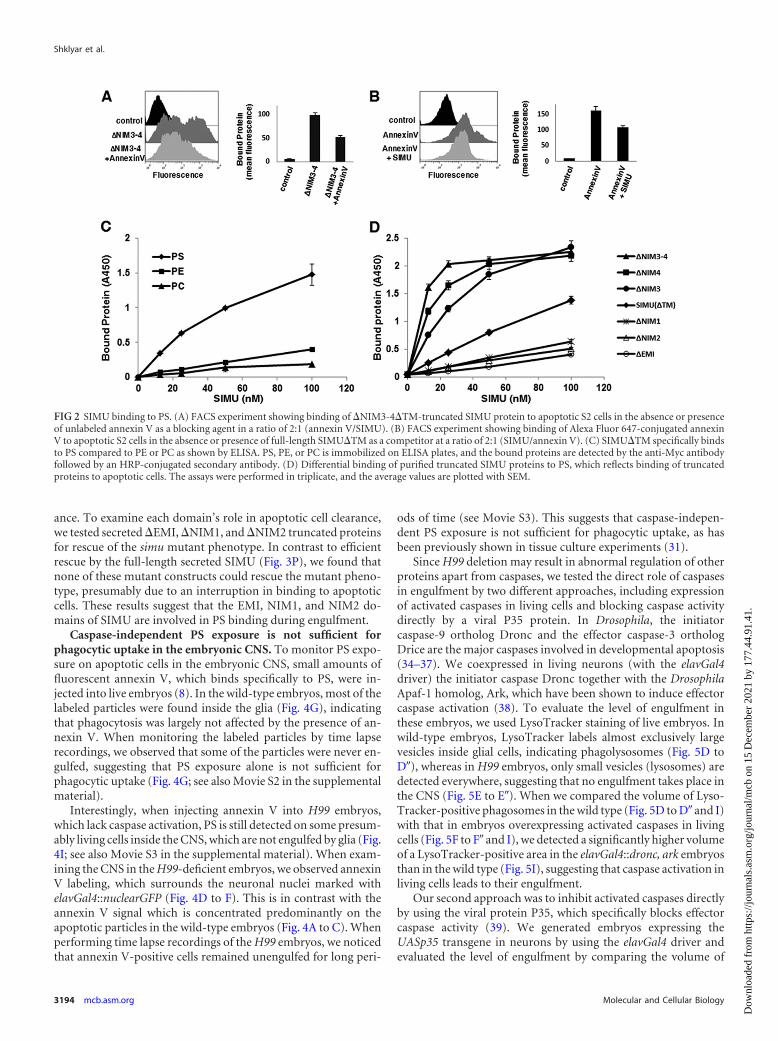
PS represents a major recognition cue for engulfment by pro-fessional and nonprofessional phagocytes (29–31). A number ofphagocytic transmembrane receptors and secreted bridging mol-ecules have been shown to bind PS (10–12, 29, 32). Moreover, PSexposure on apoptotic cells is required for phagocytic uptake andmasking PS on apoptotic surfaces inhibits engulfment. Annexin Vspecifically binds exposed PS on apoptotic cells, and it is broadlyused as a marker for apoptosis and as a PS masking agent (10, 33).In order to examine the possibility that PS is a ligand for SIMU, wetested SIMU binding to apoptotic S2 cells in the presence of an-nexin V. Binding of the �NIM3-4 mutant with strongest bindingaffinity to apoptotic S2 cells was significantly reduced by the pres-ence of annexin V compared to the strong binding of this formwith no annexin V addition (Fig. 2A), indicating that the EMI,NIM1, and NIM2 domains most likely bind PS on apoptotic cells.Moreover, in the corresponding experiment, significantly reduced
Shklyar et al.
3192 mcb.asm.org Molecular and Cellular Biology
Dow
nloa
ded
from
http
s://j
ourn
als.
asm
.org
/jour
nal/m
cb o
n 15
Dec
embe
r 20
21 b
y 17
7.44
.91.
41.

binding of fluorescently labeled annexin V to apoptotic S2 cellswas detected in the presence of SIMU in its full-length comparedto the annexin V binding with no SIMU addition (Fig. 2B). Wetherefore tested whether SIMU directly interacts with PS by usinga solid-phase binding assay for phospholipids (10). Polypropyleneplates were coated with PS, phosphatidylcholine (PC), or phos-phatidylethanolamine (PE) and incubated with purified SIMU inincreasing concentrations. According to our results, SIMU bindsspecifically to PS in a dose-dependent manner (Fig. 2C). In addi-tion, binding of SIMU truncated constructs to PS-coated plates(Fig. 2D) correlated with their binding to apoptotic cells (Fig. 1Band C), further supporting the notion that PS is an authenticSIMU ligand on apoptotic cells.
EMI, NIM1, and NIM2 are required for SIMU function invivo. Our data show that EMI, NIM1, and NIM2 are required forSIMU binding to PS on apoptotic cells in vitro. To explore the
function of these domains in vivo during embryonic CNS devel-opment, we tested the ability of the different truncated forms ofSIMU (Fig. 1A) to rescue the phenotype of simu null mutantswhen expressed specifically in glial cells under the regulation ofthe repoGal4 driver. To examine the apoptotic cell clearance phe-notype, we measured apoptotic cell volume, using an anti-acti-vated caspase 3 (CM1) antibody, in the CNS of stage 16 embryos(8) (Fig. 3B to O). As we have previously shown, in simu mutantembryos there is an accumulation of unengulfed apoptotic parti-cles and therefore an increase in apoptotic cell volume due todefects in the recognition and engulfment steps of phagocytosis(8) (Fig. 3E, F, and P). In concordance with the in vitro results, thetransmembrane �EMI-NIM2 variant, which lacks all three do-mains required for binding to PS, did not rescue the simu mutantphenotype (Fig. 3K, L, and P), implying that EMI, NIM1, andNIM2 are required in vivo for simu function in apoptotic cell clear-
FIG 1 SIMU binding to apoptotic cells. (A) Schematic of SIMU constructs designed for expression in vitro in HEK293 cells, fused to Myc and HIS tags, or fusedin vivo to GFP tags. (B and C) A representative fluorescence-activated cell sorter (FACS) experiment (left) and the quantitation graph (right) showing alteredbinding of truncated SIMU proteins to apoptotic S2 cells compared to SIMU�TM. The bound protein was detected by an anti-Myc antibody followed by afluorescently labeled secondary antibody. Note the reduced binding of truncated proteins in panel B and increased binding in panel C. The assays were performedin triplicate. Columns represent mean fluorescence values � standard errors of the means (SEM).
Caspase Role in Apoptotic Cell Clearance
August 2013 Volume 33 Number 16 mcb.asm.org 3193
Dow
nloa
ded
from
http
s://j
ourn
als.
asm
.org
/jour
nal/m
cb o
n 15
Dec
embe
r 20
21 b
y 17
7.44
.91.
41.
ance. To examine each domain’s role in apoptotic cell clearance,we tested secreted �EMI, �NIM1, and �NIM2 truncated proteinsfor rescue of the simu mutant phenotype. In contrast to efficientrescue by the full-length secreted SIMU (Fig. 3P), we found thatnone of these mutant constructs could rescue the mutant pheno-type, presumably due to an interruption in binding to apoptoticcells. These results suggest that the EMI, NIM1, and NIM2 do-mains of SIMU are involved in PS binding during engulfment.
Caspase-independent PS exposure is not sufficient forphagocytic uptake in the embryonic CNS. To monitor PS expo-sure on apoptotic cells in the embryonic CNS, small amounts offluorescent annexin V, which binds specifically to PS, were in-jected into live embryos (8). In the wild-type embryos, most of thelabeled particles were found inside the glia (Fig. 4G), indicatingthat phagocytosis was largely not affected by the presence of an-nexin V. When monitoring the labeled particles by time lapserecordings, we observed that some of the particles were never en-gulfed, suggesting that PS exposure alone is not sufficient forphagocytic uptake (Fig. 4G; see also Movie S2 in the supplementalmaterial).
Interestingly, when injecting annexin V into H99 embryos,which lack caspase activation, PS is still detected on some presum-ably living cells inside the CNS, which are not engulfed by glia (Fig.4I; see also Movie S3 in the supplemental material). When exam-ining the CNS in the H99-deficient embryos, we observed annexinV labeling, which surrounds the neuronal nuclei marked withelavGal4::nuclearGFP (Fig. 4D to F). This is in contrast with theannexin V signal which is concentrated predominantly on theapoptotic particles in the wild-type embryos (Fig. 4A to C). Whenperforming time lapse recordings of the H99 embryos, we noticedthat annexin V-positive cells remained unengulfed for long peri-
ods of time (see Movie S3). This suggests that caspase-indepen-dent PS exposure is not sufficient for phagocytic uptake, as hasbeen previously shown in tissue culture experiments (31).
Since H99 deletion may result in abnormal regulation of otherproteins apart from caspases, we tested the direct role of caspasesin engulfment by two different approaches, including expressionof activated caspases in living cells and blocking caspase activitydirectly by a viral P35 protein. In Drosophila, the initiatorcaspase-9 ortholog Dronc and the effector caspase-3 orthologDrice are the major caspases involved in developmental apoptosis(34–37). We coexpressed in living neurons (with the elavGal4driver) the initiator caspase Dronc together with the DrosophilaApaf-1 homolog, Ark, which have been shown to induce effectorcaspase activation (38). To evaluate the level of engulfment inthese embryos, we used LysoTracker staining of live embryos. Inwild-type embryos, LysoTracker labels almost exclusively largevesicles inside glial cells, indicating phagolysosomes (Fig. 5D toD�), whereas in H99 embryos, only small vesicles (lysosomes) aredetected everywhere, suggesting that no engulfment takes place inthe CNS (Fig. 5E to E�). When we compared the volume of Lyso-Tracker-positive phagosomes in the wild type (Fig. 5D to D� and I)with that in embryos overexpressing activated caspases in livingcells (Fig. 5F to F� and I), we detected a significantly higher volumeof a LysoTracker-positive area in the elavGal4::dronc, ark embryosthan in the wild type (Fig. 5I), suggesting that caspase activation inliving cells leads to their engulfment.
Our second approach was to inhibit activated caspases directlyby using the viral protein P35, which specifically blocks effectorcaspase activity (39). We generated embryos expressing theUASp35 transgene in neurons by using the elavGal4 driver andevaluated the level of engulfment by comparing the volume of
FIG 2 SIMU binding to PS. (A) FACS experiment showing binding of �NIM3-4�TM-truncated SIMU protein to apoptotic S2 cells in the absence or presenceof unlabeled annexin V as a blocking agent in a ratio of 2:1 (annexin V/SIMU). (B) FACS experiment showing binding of Alexa Fluor 647-conjugated annexinV to apoptotic S2 cells in the absence or presence of full-length SIMU�TM as a competitor at a ratio of 2:1 (SIMU/annexin V). (C) SIMU�TM specifically bindsto PS compared to PE or PC as shown by ELISA. PS, PE, or PC is immobilized on ELISA plates, and the bound proteins are detected by the anti-Myc antibodyfollowed by an HRP-conjugated secondary antibody. (D) Differential binding of purified truncated SIMU proteins to PS, which reflects binding of truncatedproteins to apoptotic cells. The assays were performed in triplicate, and the average values are plotted with SEM.
Shklyar et al.
3194 mcb.asm.org Molecular and Cellular Biology
Dow
nloa
ded
from
http
s://j
ourn
als.
asm
.org
/jour
nal/m
cb o
n 15
Dec
embe
r 20
21 b
y 17
7.44
.91.
41.

FIG 3 Phenotypic analysis of the truncated simu constructs in vivo. (A to O) Projections from confocal stacks of the CNS at embryonic stage 16, ventral view;apoptotic cells are indicated in red (CM1) and glia and macrophages in green (anti-SIMU antibody). Bar, 20 �m. (A to C) In the wild-type (wt) embryo, glia (g)and macrophages (m) are labeled with anti-SIMU in green (endogenous protein). (D to F) In the simu null mutant, no SIMU expression was detected. (G to O)In rescue experiments, SIMU expression was detected only in glial cells (repoGal4 driver). (G to I) Complete rescue of the simu null phenotype with full-lengthSIMU. (J to O) No rescue with truncated SIMU proteins expressed in glial cells. (P) Quantification of phenotypic rescue of simu null mutants by the different simutransgenes. Columns represent mean total volume of apoptotic particles within confocal stacks of the CNS � SEM (n � 7 to 10); asterisks indicate statisticalsignificance versus wild-type results, as determined by one-way analysis of variance (ANOVA). ***, P � 0.001; **, P � 0.01; n.s. (not significant), P � 0.05.
Caspase Role in Apoptotic Cell Clearance
August 2013 Volume 33 Number 16 mcb.asm.org 3195
Dow
nloa
ded
from
http
s://j
ourn
als.
asm
.org
/jour
nal/m
cb o
n 15
Dec
embe
r 20
21 b
y 17
7.44
.91.
41.

FIG 4 Dynamic analysis of PS exposure in the embryonic CNS. (A to F) Nuclear GFP driven with elavGal4 labels neuronal nuclei of wild-type (A and C) and H99(D and F) embryonic CNS. Bar, 20 �m. (B, C, E, and F) PS on neuronal membranes is marked by annexin V in red. Arrows depict selected neurons with annexinV on their membranes. (G to I) Time-lapse recordings of phagocytosis in stage 16 embryos. Glia and macrophages are labeled with simu-cytGFP (green), and PSexposure is labeled by the fluorescent annexin V (red); selected frames are shown (movies are available in the supplemental material). (G) In the wild type, mostof the PS-positive particles are inside the glial cells. (H) An engulfment event is marked. Some annexin V-positive particles remained unengulfed for long periodsof time (arrows). (I) H99-deficient embryonic CNS with annexin V-positive particles remaining outside the glia and not engulfed.
Shklyar et al.
3196 mcb.asm.org Molecular and Cellular Biology
Dow
nloa
ded
from
http
s://j
ourn
als.
asm
.org
/jour
nal/m
cb o
n 15
Dec
embe
r 20
21 b
y 17
7.44
.91.
41.

LysoTracker-positive phagosomes in wild-type embryos to that inthe elavGal4::p35 embryos. Our results show that, consistent withthe reduced volume of the CM1-positive particles in the elavGal4::p35 embryos (Fig. 5C to C� and H), there was a lower volume ofLysoTracker-positive area compared to the volume in the wildtype (Fig. 5G to G� and I), indicating that inhibition of effectorcaspases leads to a decrease in engulfment. Overall, these resultssuggest that caspase activity is required for engulfment of apop-totic cells. Therefore, we propose that in addition to PS exposureon apoptotic surfaces, other ligands which are caspase dependentare required for apoptotic cell clearance.
SIMU is localized in patches on phagocytic membranes dur-ing engulfment. To follow SIMU-PS interactions during clear-ance of apoptotic neurons, we examined SIMU expression and
distribution on glial membranes using an anti-SIMU antibody.We noticed that SIMU is not homogeneously distributed onphagocytic glial membranes. The patchy distribution of SIMUsuggests clustering of the protein (Fig. 5A and A�). To test whetherthis specific pattern correlates with engulfment, we exploredSIMU localization in H99-deficient embryos, in which no phago-cytic activity of glia can be detected, using LysoTracker staining oflive embryos (Fig. 5E= and E�). When we examined SIMU expres-sion in these embryos, we found homogeneous rather than patchydistribution of the protein on the phagocytic membranes (Fig. 5Band B�). This result suggests that SIMU forms clusters only duringengulfment, possibly by interacting with its putative partner(s) onthe phagocytic membranes, consistent with a “tethering and tick-ling” multiple-receptor clustering model (7). In H99 mutant em-
FIG 5 Caspase activity affects SIMU localization and phagocytosis of apoptotic cells. (A to C) Projections from confocal stacks of the CNS at embryonic stage16. A ventral view with apoptotic particles labeled with CM1 (red) and glial cells labeled with anti-SIMU (green) is shown. Bar, 20 �m. (A to A�) Wild type. SIMUis localized in patches on glial membranes (arrows). (B to B�) H99 mutant embryo with no CM1 staining (B=) and SIMU is localized homogeneously on glialmembranes (B and B�). (C to C� and H) elavGal4::p35 embryos with much lower CM1 staining (C=, C�, H). (D to F) Projections from confocal stacks of the CNSat embryonic stage 16. A ventral view with phagosomes and lysosomes labeled with LysoTracker (red) and glial cells and macrophages labeled with simu-cytGFP(green) is shown. (D to D�) Wild type. (E to E�) H99. (F to F�) elavGal4::dronc, ark. (G to G�) elavGal4::p35. (H and I) Columns represent mean total volume ofapoptotic particles (H) or LysoTracker-positive area (I) within confocal stacks of the CNS � SEM (n � 7 to 10); asterisks indicate statistical significance versuswild-type results, as determined by one-way ANOVA. ***, P � 0.001; **, P � 0.01.
Caspase Role in Apoptotic Cell Clearance
August 2013 Volume 33 Number 16 mcb.asm.org 3197
Dow
nloa
ded
from
http
s://j
ourn
als.
asm
.org
/jour
nal/m
cb o
n 15
Dec
embe
r 20
21 b
y 17
7.44
.91.
41.
bryos, inhibition of initiator and effector caspases takes place, asconfirmed by a lack of staining with CM1 antibody (Fig. 5Band B�).
To address the important issue of whether ligand binding isneeded for receptor clustering, we examined SIMU localization inembryos expressing the �EMI-NIM2 SIMU form, which does notbind to PS. In these embryos, no SIMU patches were detected onglial membranes, suggesting that initial binding of SIMU to PS isrequired for its clustering with other factors (Fig. 3J).
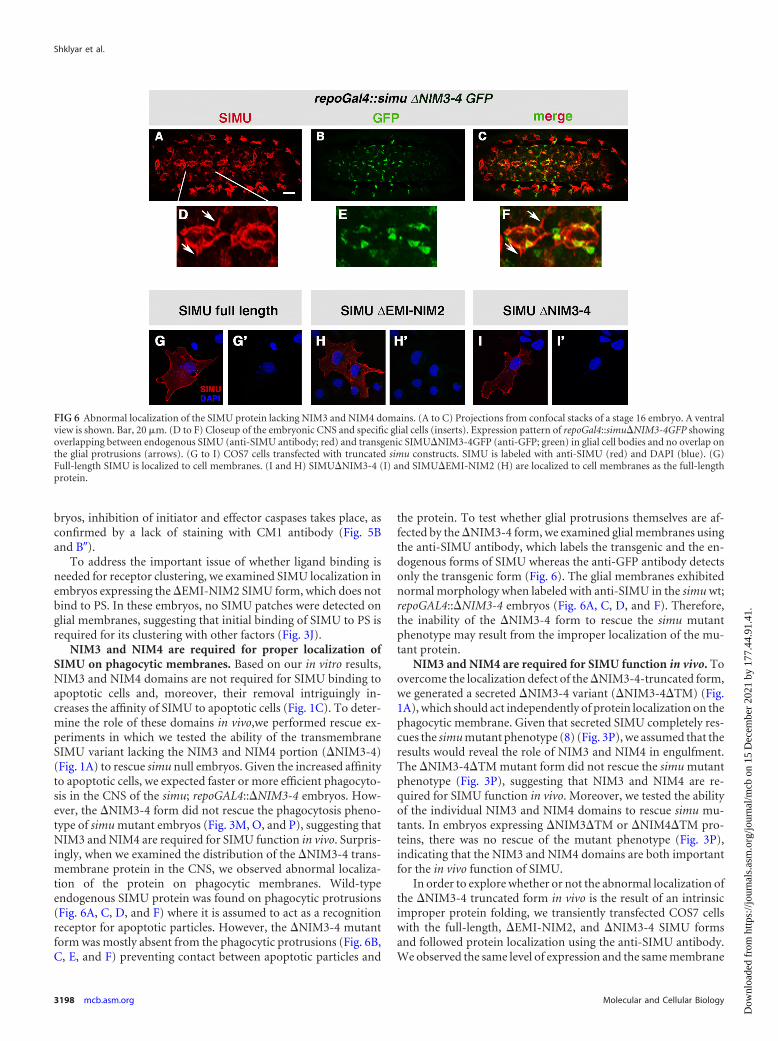
NIM3 and NIM4 are required for proper localization ofSIMU on phagocytic membranes. Based on our in vitro results,NIM3 and NIM4 domains are not required for SIMU binding toapoptotic cells and, moreover, their removal intriguingly in-creases the affinity of SIMU to apoptotic cells (Fig. 1C). To deter-mine the role of these domains in vivo,we performed rescue ex-periments in which we tested the ability of the transmembraneSIMU variant lacking the NIM3 and NIM4 portion (�NIM3-4)(Fig. 1A) to rescue simu null embryos. Given the increased affinityto apoptotic cells, we expected faster or more efficient phagocyto-sis in the CNS of the simu; repoGAL4::�NIM3-4 embryos. How-ever, the �NIM3-4 form did not rescue the phagocytosis pheno-type of simu mutant embryos (Fig. 3M, O, and P), suggesting thatNIM3 and NIM4 are required for SIMU function in vivo. Surpris-ingly, when we examined the distribution of the �NIM3-4 trans-membrane protein in the CNS, we observed abnormal localiza-tion of the protein on phagocytic membranes. Wild-typeendogenous SIMU protein was found on phagocytic protrusions(Fig. 6A, C, D, and F) where it is assumed to act as a recognitionreceptor for apoptotic particles. However, the �NIM3-4 mutantform was mostly absent from the phagocytic protrusions (Fig. 6B,C, E, and F) preventing contact between apoptotic particles and
the protein. To test whether glial protrusions themselves are af-fected by the �NIM3-4 form, we examined glial membranes usingthe anti-SIMU antibody, which labels the transgenic and the en-dogenous forms of SIMU whereas the anti-GFP antibody detectsonly the transgenic form (Fig. 6). The glial membranes exhibitednormal morphology when labeled with anti-SIMU in the simu wt;repoGAL4::�NIM3-4 embryos (Fig. 6A, C, D, and F). Therefore,the inability of the �NIM3-4 form to rescue the simu mutantphenotype may result from the improper localization of the mu-tant protein.
NIM3 and NIM4 are required for SIMU function in vivo. Toovercome the localization defect of the �NIM3-4-truncated form,we generated a secreted �NIM3-4 variant (�NIM3-4�TM) (Fig.1A), which should act independently of protein localization on thephagocytic membrane. Given that secreted SIMU completely res-cues the simu mutant phenotype (8) (Fig. 3P), we assumed that theresults would reveal the role of NIM3 and NIM4 in engulfment.The �NIM3-4�TM mutant form did not rescue the simu mutantphenotype (Fig. 3P), suggesting that NIM3 and NIM4 are re-quired for SIMU function in vivo. Moreover, we tested the abilityof the individual NIM3 and NIM4 domains to rescue simu mu-tants. In embryos expressing �NIM3�TM or �NIM4�TM pro-teins, there was no rescue of the mutant phenotype (Fig. 3P),indicating that the NIM3 and NIM4 domains are both importantfor the in vivo function of SIMU.
In order to explore whether or not the abnormal localization ofthe �NIM3-4 truncated form in vivo is the result of an intrinsicimproper protein folding, we transiently transfected COS7 cellswith the full-length, �EMI-NIM2, and �NIM3-4 SIMU formsand followed protein localization using the anti-SIMU antibody.We observed the same level of expression and the same membrane
FIG 6 Abnormal localization of the SIMU protein lacking NIM3 and NIM4 domains. (A to C) Projections from confocal stacks of a stage 16 embryo. A ventralview is shown. Bar, 20 �m. (D to F) Closeup of the embryonic CNS and specific glial cells (inserts). Expression pattern of repoGal4::simu�NIM3-4GFP showingoverlapping between endogenous SIMU (anti-SIMU antibody; red) and transgenic SIMU�NIM3-4GFP (anti-GFP; green) in glial cell bodies and no overlap onthe glial protrusions (arrows). (G to I) COS7 cells transfected with truncated simu constructs. SIMU is labeled with anti-SIMU (red) and DAPI (blue). (G)Full-length SIMU is localized to cell membranes. (I and H) SIMU�NIM3-4 (I) and SIMU�EMI-NIM2 (H) are localized to cell membranes as the full-lengthprotein.
Shklyar et al.
3198 mcb.asm.org Molecular and Cellular Biology
Dow
nloa
ded
from
http
s://j
ourn
als.
asm
.org
/jour
nal/m
cb o
n 15
Dec
embe
r 20
21 b
y 17
7.44
.91.
41.

localization of all the forms (Fig. 6G to I=), suggesting that theabnormal distribution of �NIM3-4 form in vivo may result fromthe impaired interaction with additional factors on the plasmamembrane.
DISCUSSION
In this study, we have shown that caspase activity is needed forefficient and precise phagocytosis of apoptotic particles duringembryonic CNS development. Focusing on the recognition recep-tor SIMU, we have demonstrated that caspase-independent PSexposure is not sufficient for clearance. Since there are no known“don’t eat me” signals in Drosophila, this mechanism may explainwhy living cells exposing (transiently or in small amounts) some“eat me” signals on their surfaces are not removed until they areactually dying.
PS is the most abundant “eat me” signal (40, 41) and is recog-nized directly by different receptors in mammals (29, 42), includ-ing the TIM family (12, 43, 44), BAI1 (45), and SIMU homologstabilin-2 (13), and by DRPR in Drosophila (46). Here we demon-strate that the fly phagocytic receptor SIMU recognizes PS onapoptotic cell surfaces, illustrating once more the high level ofevolutionary conservation of apoptotic cell clearance between in-sects and vertebrates. Moreover, we show that SIMU specificallybinds to PS on apoptotic cells in vitro through its EMI, NIM1, andNIM2 domains and that removal of these domains affects SIMUfunction in vivo and its distribution on glial membranes. In theembryonic CNS, SIMU protein is localized in patches on glialphagocytic membranes, suggesting clustering of SIMU with itselfor other phagocytic receptors. This distribution depends on theinitial binding of SIMU to PS, since the truncated �EMI-NIM2SIMU does not make patches on the membranes.
Receptor clustering is thought to be important for proper andefficient removal of apoptotic particles (29). The “tethering andtickling” model (7) speaks about multiple interactions betweenphagocytic receptors and their ligands, which result in receptorclustering and signaling for engulfment. Elucidation of the molec-ular mechanisms underlying receptor clustering remains elusive,and it is unknown what the interactions are between differentphagocytic receptors and whether they are influenced by ligandbinding and conformation changes. In H99 embryos, lackingcaspase activation, where no engulfment takes place, SIMU isequally distributed on glial membranes, indicating that SIMUclusters with itself or other receptors only during engulfment.Based on our structure-function analysis, we show that SIMUclustering during engulfment depends on its binding to PS, sug-gesting that clustering of phagocytic receptors depends on theirbinding to the ligands. What are these ligands on apoptotic cells,which are absent in H99 mutants?
Caspase involvement in the expression and exposure of “findme” and “eat me” signals is not completely understood. Particu-larly, PS exposure on apoptotic cells is considered caspase depen-dent in worms and mammals and is required for engulfment ofapoptotic particles. Moreover, recent discoveries from S. Nagata’slaboratory showed that PS exposure on living cells is not sufficientfor their phagocytic removal in vitro (31). However, the study byvan Delft and colleagues in Casp9/ mice showed that intactslowly dying Casp9/ mutant thymocytes exposed PS withoutcaspases, which led to their noninflammatory phagocytosis (47).To test how caspase activity affects PS exposure and apoptotic cellclearance in Drosophila, we examined H99 mutant embryos,
which lack caspase activation. In these embryos, we detected PSaccumulation on some presumably living cells. Following thesecells by time lapse recording, we observed that they were neverengulfed, indicating that PS accumulation on their surface is notsufficient for clearance. Moreover, in H99 embryos, SIMU is ex-pressed on phagocytic membranes and its ligand PS is detected onsome cell surfaces but no engulfment takes place, implying thatsomething is missing. We suggest that additional caspase-depen-dent ligands on apoptotic particles are missing which are recog-nized by their receptors on phagocytic membranes and that, akinto the “tethering and tickling” model, they participate in receptorclustering leading to engulfment. Finding these additionalcaspase-dependent ligands will help us uncover the molecular ba-sis of multiple receptor/ligand interactions.
Interestingly, based on the structure-function analysis ofSIMU, we discovered a novel internal inhibitory mechanism inthe SIMU receptor which presumably prevents engulfment of liv-ing cells expressing PS on their surface. We found that the SIMUNIM3 and NIM4 domains are not involved in PS recognition andbinding but rather inhibit SIMU binding to PS. Furthermore,these domains are required for SIMU proper localization andfunction in vivo in apoptotic cell clearance, suggesting their role inengulfment. We propose that they may be involved in interactionsof SIMU with other phagocytic receptors or bridging molecules,which recognize additional caspase-dependent and/or -indepen-dent alterations on apoptotic surfaces. These interactions throughthe NIM3 and NIM4 domains may change SIMU conformation,relieve the internal inhibition, and dramatically increase SIMUaffinity to PS.
Taken together, our data suggest the following model forSIMU action in apoptotic cell clearance (Fig. 7). SIMU recognizesPS on apoptotic cells with mild affinity that is crucial for not en-gulfing living cells, which have been shown to expose (transientlyor in small amounts) PS on their surfaces (48, 49). When addi-tional “eat me” signals are exposed on apoptotic cells, indicatingthat the cells are actually dying, and are recognized by additionalreceptors (putative SIMU partners), SIMU affinity to PS dramat-ically increases. We propose that this increase in affinity occurs asa result of relieving the internal inhibition by NIM3 and NIM4through binding of the partner(s) to SIMU and changing its con-formation.
Observing the phagocytic behavior of glia in the wild-type em-bryonic CNS, we found that glial protrusions probe the environ-ment before engulfing an apoptotic cell (see Movie S4 in the sup-
FIG 7 PS and caspase activation are required for engulfment. A schematicrepresentation is shown of PS and additional caspase-dependent ligands onapoptotic cells that are recognized by different receptors on the phagocyticsurface, which cluster and thus lead to engulfment of the apoptotic cell by thephagocyte.
Caspase Role in Apoptotic Cell Clearance
August 2013 Volume 33 Number 16 mcb.asm.org 3199
Dow
nloa
ded
from
http
s://j
ourn
als.
asm
.org
/jour
nal/m
cb o
n 15
Dec
embe
r 20
21 b
y 17
7.44
.91.
41.

plemental material). Instances of phagocytes reaching theapoptotic cell without uptake suggest a threshold of receptor af-finity which has to be reached and/or involvement of additionalfactors that are needed to be active for proper engulfment. Thismodel suggests a mechanism for controlling the removal of onlydying and not healthy cells, which is crucial for normal develop-ment and homeostasis of multicellular organisms. Finding theadditional caspase-dependent ligands and understanding howapoptotic cells are marked for accurate and efficient eliminationwill provide new insight into the molecular and cellular mecha-nisms of apoptotic cell clearance.
ACKNOWLEDGMENTS
We thank V. Auld, O. Shuldiner, H. Steller, E. Arama, and the Blooming-ton Stock Center for generously providing fly strains. We thank A. Salz-berg and T. Schultheiss for comments on the manuscript and the Kurantlaboratory members for support and constructive criticism.
This work was supported by grants from IRG (grant no. 2498084), theIsrael Ministry of Health (grant no. 3-00000-6162), and the Israel ScienceFoundation (grant no. 427/11).
REFERENCES1. Kinchen JM, Doukoumetzidis K, Almendinger J, Stergiou L, Tosello-
Trampont A, Sifri CD, Hengartner MO, Ravichandran KS. 2008. Apathway for phagosome maturation during engulfment of apoptotic cells.Nat. Cell Biol. 10:556 –566.
2. Kinchen JM, Ravichandran KS. 2007. Journey to the grave: signalingevents regulating removal of apoptotic cells. J. Cell Sci. 120:2143–2149.
3. Lauber K, Blumenthal SG, Waibel M, Wesselborg S. 2004. Clearance ofapoptotic cells: getting rid of the corpses. Mol. Cell 14:277–287.
4. Ravichandran KS. 2011. Beginnings of a good apoptotic meal: the find-me and eat-me signaling pathways. Immunity 35:445– 455.
5. Ravichandran KS, Lorenz U. 2007. Engulfment of apoptotic cells: signalsfor a good meal. Nat. Rev. Immunol. 7:964 –974.
6. Stuart LM, Ezekowitz RA. 2005. Phagocytosis: elegant complexity. Im-munity 22:539 –550.
7. Henson PM, Hume DA. 2006. Apoptotic cell removal in developmentand tissue homeostasis. Trends Immunol. 27:244 –250.
8. Kurant E, Axelrod S, Leaman D, Gaul U. 2008. Six-microns-under actsupstream of Draper in the glial phagocytosis of apoptotic neurons. Cell133:498 –509.
9. Wu HH, Bellmunt E, Scheib JL, Venegas V, Burkert C, Reichardt LF,Zhou Z, Farinas I, Carter BD. 2009. Glial precursors clear sensory neu-ron corpses during development via Jedi-1, an engulfment receptor. Nat.Neurosci. 12:1534 –1541.
10. Hanayama R, Tanaka M, Miwa K, Shinohara A, Iwamatsu A, Nagata S.2002. Identification of a factor that links apoptotic cells to phagocytes.Nature 417:182–187.
11. Kim S, Park SY, Kim SY, Bae DJ, Pyo JH, Hong M, Kim IS. 2012. Crosstalk between engulfment receptors stabilin-2 and integrin alphavbeta5orchestrates engulfment of phosphatidylserine-exposed erythrocytes.Mol. Cell. Biol. 32:2698 –2708.
12. Miyanishi M, Tada K, Koike M, Uchiyama Y, Kitamura T, Nagata S.2007. Identification of Tim4 as a phosphatidylserine receptor. Nature 450:435– 439.
13. Park SY, Jung MY, Kim HJ, Lee SJ, Kim SY, Lee BH, Kwon TH, ParkRW, Kim IS. 2008. Rapid cell corpse clearance by stabilin-2, a membranephosphatidylserine receptor. Cell Death Differ. 15:192–201.
14. Park SY, Jung MY, Lee SJ, Kang KB, Gratchev A, Riabov V, Kzhysh-kowska J, Kim IS. 2009. Stabilin-1 mediates phosphatidylserine-dependent clearance of cell corpses in alternatively activated macro-phages. J. Cell Sci. 122:3365–3373.
15. Franc NC, Heitzler P, Ezekowitz RA, White K. 1999. Requirement forcroquemort in phagocytosis of apoptotic cells in Drosophila. Science 284:1991–1994.
16. Freeman MR, Delrow J, Kim J, Johnson E, Doe CQ. 2003. Unwrappingglial biology: Gcm target genes regulating glial development, diversifica-tion, and function. Neuron 38:567–580.
17. Kurucz E, Markus R, Zsamboki J, Folkl-Medzihradszky K, Darula Z,
Vilmos P, Udvardy A, Krausz I, Lukacsovich T, Gateff E, Zettervall CJ,Hultmark D, Ando I. 2007. Nimrod, a putative phagocytosis receptorwith EGF repeats in Drosophila plasmatocytes. Curr. Biol. 17:649 – 654.
18. Somogyi K, Sipos B, Penzes Z, Kurucz E, Zsamboki J, Hultmark D,Ando I. 2008. Evolution of genes and repeats in the Nimrod superfamily.Mol. Biol. Evol. 25:2337–2347.
19. Hashimoto Y, Tabuchi Y, Sakurai K, Kutsuna M, Kurokawa K, AwasakiT, Sekimizu K, Nakanishi Y, Shiratsuchi A. 2009. Identification oflipoteichoic acid as a ligand for draper in the phagocytosis of Staphylococ-cus aureus by Drosophila hemocytes. J. Immunol. 183:7451–7460.
20. Kocks C, Cho JH, Nehme N, Ulvila J, Pearson AM, Meister M, StromC, Conto SL, Hetru C, Stuart LM, Stehle T, Hoffmann JA, ReichhartJM, Ferrandon D, Ramet M, Ezekowitz RA. 2005. Eater, a transmem-brane protein mediating phagocytosis of bacterial pathogens in Drosoph-ila. Cell 123:335–346.
21. Manaka J, Kuraishi T, Shiratsuchi A, Nakai Y, Higashida H, Henson P,Nakanishi Y. 2004. Draper-mediated and phosphatidylserine-independent phagocytosis of apoptotic cells by Drosophila hemocytes/macrophages. J. Biol. Chem. 279:48466 – 48476.
22. Thornberry NA, Lazebnik Y. 1998. Caspases: enemies within. Science281:1312–1316.
23. Yuan J, Shaham S, Ledoux S, Ellis HM, Horvitz HR. 1993. The C.elegans cell death gene ced-3 encodes a protein similar to mammalianinterleukin-1 beta-converting enzyme. Cell 75:641– 652.
24. Riedl SJ, Shi Y. 2004. Molecular mechanisms of caspase regulation duringapoptosis. Nat. Rev. Mol. Cell Biol. 5:897–907.
25. Salvesen GS, Riedl SJ. 2008. Caspase mechanisms. Adv. Exp. Med. Biol.615:13–23.
26. White K, Grether ME, Abrams JM, Young L, Farrell K, Steller H. 1994.Genetic control of programmed cell death in Drosophila. Science 264:677– 683.
27. Venken KJ, He Y, Hoskins RA, Bellen HJ. 2006. P[acman]: a BACtransgenic platform for targeted insertion of large DNA fragments in D.melanogaster. Science 314:1747–1751.
28. Thur J, Rosenberg K, Nitsche DP, Pihlajamaa T, Ala-Kokko L, Heine-gard D, Paulsson M, Maurer P. 2001. Mutations in cartilage oligomericmatrix protein causing pseudoachondroplasia and multiple epiphysealdysplasia affect binding of calcium and collagen I, II, and IX. J. Biol. Chem.276:6083– 6092.
29. Ravichandran KS. 2010. Find-me and eat-me signals in apoptotic cellclearance: progress and conundrums. J. Exp. Med. 207:1807–1817.
30. Rosenbaum S, Kreft S, Etich J, Frie C, Stermann J, Grskovic I, Frey B,Mielenz D, Poschl E, Gaipl U, Paulsson M, Brachvogel B. 2011. Iden-tification of novel binding partners (annexins) for the cell death signalphosphatidylserine and definition of their recognition motif. J. Biol.Chem. 286:5708 –5716.
31. Segawa K, Suzuki J, Nagata S. 2011. Constitutive exposure of phospha-tidylserine on viable cells. Proc. Natl. Acad. Sci. U. S. A. 108:19246 –19251.
32. Anderson HA, Maylock CA, Williams JA, Paweletz CP, Shu H, ShacterE. 2003. Serum-derived protein S binds to phosphatidylserine and stim-ulates the phagocytosis of apoptotic cells. Nat. Immunol. 4:87–91.
33. van den Eijnde SM, Boshart L, Baehrecke EH, De Zeeuw CI, Reute-lingsperger CP, Vermeij-Keers C. 1998. Cell surface exposure of phos-phatidylserine during apoptosis is phylogenetically conserved. Apoptosis3:9 –16.
34. Cooper DM, Granville DJ, Lowenberger C. 2009. The insect caspases.Apoptosis 14:247–256.
35. Kumar S. 2007. Caspase function in programmed cell death. Cell DeathDiffer. 14:32– 43.
36. Steller H. 2008. Regulation of apoptosis in Drosophila. Cell Death Differ.15:1132–1138.
37. Xu D, Woodfield SE, Lee TV, Fan Y, Antonio C, Bergmann A. 2009.Genetic control of programmed cell death (apoptosis) in Drosophila. Fly(Austin) 3:78 –90.
38. Florentin A, Arama E. 2012. Caspase levels and execution efficienciesdetermine the apoptotic potential of the cell. J. Cell Biol. 196:513–527.
39. Hay BA, Wolff T, Rubin GM. 1994. Expression of baculovirus P35prevents cell death in Drosophila. Development 120:2121–2129.
40. Fadok VA, Bratton DL, Frasch SC, Warner ML, Henson PM. 1998. Therole of phosphatidylserine in recognition of apoptotic cells by phagocytes.Cell Death Differ. 5:551–562.
41. Fadok VA, Voelker DR, Campbell PA, Cohen JJ, Bratton DL, HensonPM. 1992. Exposure of phosphatidylserine on the surface of apoptotic
Shklyar et al.
3200 mcb.asm.org Molecular and Cellular Biology
Dow
nloa
ded
from
http
s://j
ourn
als.
asm
.org
/jour
nal/m
cb o
n 15
Dec
embe
r 20
21 b
y 17
7.44
.91.
41.

lymphocytes triggers specific recognition and removal by macrophages. J.Immunol. 148:2207–2216.
42. Bratton DL, Henson PM. 2008. Apoptotic cell recognition: will the realphosphatidylserine receptor(s) please stand up? Curr. Biol. 18:R76 –R79.
43. Kobayashi N, Karisola P, Pena-Cruz V, Dorfman DM, Jinushi M,Umetsu SE, Butte MJ, Nagumo H, Chernova I, Zhu B, Sharpe AH,Ito S, Dranoff G, Kaplan GG, Casasnovas JM, Umetsu DT, DekruyffRH, Freeman GJ. 2007. TIM-1 and TIM-4 glycoproteins bind phos-phatidylserine and mediate uptake of apoptotic cells. Immunity 27:927–940.
44. Santiago C, Ballesteros A, Tami C, Martinez-Munoz L, Kaplan GG,Casasnovas JM. 2007. Structures of T cell immunoglobulin mucin recep-tors 1 and 2 reveal mechanisms for regulation of immune responses by theTIM receptor family. Immunity 26:299 –310.
45. Park D, Tosello-Trampont AC, Elliott MR, Lu M, Haney LB, Ma Z,Klibanov AL, Mandell JW, Ravichandran KS. 2007. BAI1 is an engulf-
ment receptor for apoptotic cells upstream of the ELMO/Dock180/Racmodule. Nature 450:430 – 434.
46. Tung TT, Nagaosa K, Fujita Y, Kita A, Mori H, Okada R, Nonaka S,Nakanishi Y. 2013. Phosphatidylserine recognition and induction ofapoptotic cell clearance by Drosophila engulfment receptor Draper. J.Biochem. 153:483– 491.
47. van Delft MF, Smith DP, Lahoud MH, Huang DC, Adams JM. 2010.Apoptosis and non-inflammatory phagocytosis can be induced by mito-chondrial damage without caspases. Cell Death Differ. 17:821– 832.
48. Dillon SR, Mancini M, Rosen A, Schlissel MS. 2000. Annexin V binds toviable B cells and colocalizes with a marker of lipid rafts upon B cellreceptor activation. J. Immunol. 164:1322–1332.
49. Hamon Y, Broccardo C, Chambenoit O, Luciani MF, Toti F, Chaslin S,Freyssinet JM, Devaux PF, McNeish J, Marguet D, Chimini G. 2000.ABC1 promotes engulfment of apoptotic cells and transbilayer redistribu-tion of phosphatidylserine. Nat. Cell Biol. 2:399 – 406.
Caspase Role in Apoptotic Cell Clearance
August 2013 Volume 33 Number 16 mcb.asm.org 3201
Dow
nloa
ded
from
http
s://j
ourn
als.
asm
.org
/jour
nal/m
cb o
n 15
Dec
embe
r 20
21 b
y 17
7.44
.91.
41.